Capítulo 3
CUIDAR DE LOS DEMÁS Y APRECIARLOS
¿Qué es lo que ocurre en el cerebro para que un animal cuide de los demás o exprese valores sociales? Según la hipótesis predominante, esto se explica básicamente por la neuroquímica del apego y los vínculos afectivos de los mamíferos.1 Por tanto, con el fin de entender la plataforma cerebral de los valores sociales, primero tenemos que plantearnos la pregunta más fundamental, que nos conducirá de vuelta a los valores sociales: ¿cómo es posible que los cerebros se preocupen por algo? O, dicho de otro modo, ¿cómo es posible que las neuronas otorguen valor a algo?
La primera parte de esta historia, y la más importante, tiene que ver con la autopreservación.2 Todos los sistemas nerviosos se organizan para cuidar de la supervivencia básica del cuerpo del que forman parte. Desde una perspectiva evolutiva, lo fundamental es muy sencillo: se elige el «cuidado» de uno mismo frente al «descuido» de uno mismo. Los animales que no consiguen preservarse a sí mismos no tienen posibilidad alguna de transmitir sus genes, mientras que los animales que consiguen mantener sanos sus cuerpos tienen opciones para transmitir sus genes. Para que un animal sobreviva, se pone el mundo patas arriba para conseguir energía, agua y todo lo necesario para que el organismo funcione. El dolor y el miedo son señales de supervivencia que indican la necesidad de una conducta correctiva. Los distintos tipos de dolor indican distintas vías para la corrección de una conducta.
Estas observaciones de carácter general suscitan preguntas sobre los mecanismos neuronales: ¿cómo es posible que un ratón, por ejemplo, sepa el lugar donde encontrar comida, se apresure a entrar en una madriguera o construya una ratonera?; ¿de qué modo las decisiones conductuales que sirven para conseguir nuestro bienestar llegan a las neuronas?
La respuesta más sencilla a esta pregunta es que las neuronas del tallo cerebral y del hipotálamo del ratón controlan el «medio interno» del ratón; es decir, el estado interior de su cuerpo en relación a los parámetros que son importantes para la supervivencia. Cuando se detecta una necesidad en concreto, se genera una emoción motivacional. En el ratón, del mismo modo que en los humanos, el tallo cerebral y las neuronas hipotalámicas regulan la temperatura corporal, los niveles de glucosa, la presión sanguínea, el ritmo cardíaco y los niveles de dióxido de carbono. La homeostasis es el proceso mediante el cual el medio interno del organismo se regula para aproximarse al rango necesario para la supervivencia. Y el dolor, tal como ha observado el neurocientífico Bud Craig, es una emoción homeostática.3 Todos nosotros estamos familiarizados con los cambios del medio interno que indican la necesidad de corregir un desequilibrio: el pánico que experimentamos cuando nos falta el oxígeno; la incomodidad que sentimos cuando tenemos frío; la sensación de sed, las náuseas y el dolor que produce el hambre extrema. Estas señales van acompañadas de distintos impulsos: el de buscar calor, agua, alimento, vomitar, escaparse, acurrucarse, etcétera.
Al utilizar señales perceptuales, como los olores y los sonidos, el cerebro subcortical del ratón también hace una valoración de los riesgos y las oportunidades que ofrece el mundo exterior. En las ratoncitas, el olor de las semillas provoca una conducta de acercamiento; en los ratoncitos, el olor que desprende una hembra en celo genera una conducta de cortejo. Un ratón que se adentre en un territorio nuevo en el que huela el orín de otro ratón macho se marchará a otra parte.
En nuestro caso, el miedo que suscita en nosotros el gruñir de un perro o el pánico inducido por un humo impredecible son sensaciones inconfundibles y desagradables. Estas sensaciones importantísimas para preservar la vida se integran en el organismo, y se coordinan con un movimiento de respuesta adecuado por parte de las estructuras subcorticales del tallo cerebral y del hipotálamo, así como en la corteza insular y la corteza cingulada (véase figura 3.1). Los mecanismos del sistema nervioso simpático preparan el cuerpo para «la lucha o la huida», y cuando la amenaza ha pasado, otros mecanismos del sistema parasimpático restablecen la presión sanguínea y el ritmo cardíaco a un estado de menor desgaste energético que nos invita a «descansar y digerir». Además, el circuito es sensible a las prioridades, de modo que el miedo a un depredador que se acerque supera a las ganas de comer una sabrosa nuez o al deseo de aparearse con una hembra en celo.
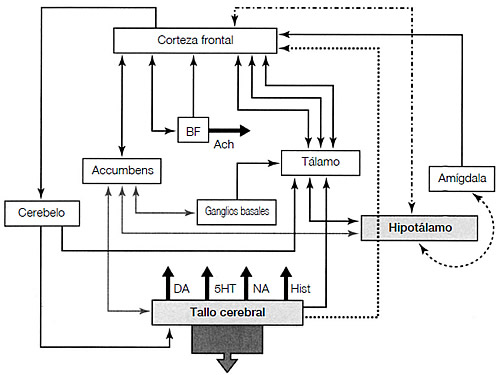
FIGURA 3.1. Esta figura ilustra esquemáticamente algunas de las estructuras subcorticales y su relación con la corteza cerebral. Obsérvense, en concreto, las numerosas vías que existen entre las zonas frontales y las subcorticales, incluidas las que implican una recompensa (el núcleo accumbens) y el miedo (amígdala). El hipotálamo está ampliamente relacionado con muchas estructuras, y suele estarlo de forma bidireccional. Del tallo cerebral surgen cuatro sistemas de proyección neuronal con sustancias neuroquímicas distintas, y cada sistema llega a muchas zonas distintas. Las cuatro sustancias neuroquímicas, que a veces se denominan neuromoduladores, son la serotonina (5HT), la noradrenalina (NA), la dopamina (DA) y la histamina (Hist). Diagrama basado en los datos de Josef Parvizi, «Corticocentric Myopia: Old Bias in New Cognitive Sciences», Trends in Cognitive Sciences 13, nº 8, 2009, págs. 354-359. (Para ver la ubicación de la amígdala en un espacio tridimensional, véase <http:// commons.wikimedia.org/wiki/File:Amygdala_small.gif>.)
El circuito límbico del tallo cerebral, al integrar las señales tanto del medio interno como de la superficie corporal, supone la base organizativa que se encarga de la autopreservación, y, por consiguiente, de un mínimo sentido del ser.4 Conservar la salud y el bienestar del cuerpo constituye el andamiaje neurobiológico para los niveles elevados de autorrepresentación, como la sensación que se tiene de uno mismo como persona que pertenece a un grupo social y que tiene vínculos especiales con algunos individuos concretos.5
Así pues, en un sentido muy elemental, el «cuidado» es una función básica de los sistemas nerviosos. Los cerebros están organizados para procurar el bienestar y para buscar alivio ante cualquier incomodidad. Por tanto, de un modo perfectamente claro y directo, el circuito para la autopreservación y el rechazo del dolor se encuentra en el origen de los valores más elementales: los valores de permanecer vivo y bien. Para las ranas, los salmones y los tritones, este tipo de cuidado es todo lo que tienen, y poco más. No obstante, este nivel de integración, que todos los seres vertebrados tienen en alta consideración, es de una complejidad exquisita.
La presión que ejerce el proceso de selección para procurarse el cuidado de uno mismo es evidente, aun cuando el circuito subyacente se vea oscurecido por su propia complejidad. ¿Cómo podríamos explicar el cuidado o la atención a los demás? Tal y como propuse en el capítulo 2, la idea fundamental es que, en el caso de los mamíferos, los ajustes evolutivos en los sistemas emocionales, endocrino, nervioso y de recompensa/ castigo amplían de un modo efectivo el rango de individuos por los que el animal se preocupa, al menos en lo relativo a un conjunto determinado de conductas relevantes para la supervivencia. De este modo, la madre ratón se comporta como si sus recién nacidos estuvieran incluidos en su ámbito básico homeostático: se les debe alimentar, limpiar y calentar, y también hay que protegerlos de los distintos peligros del mundo. Cuando los cachorros están amenazados, su bienestar le importa a la hembra tanto como su propio bienestar, y por consiguiente es capaz de emprender acciones correctivas. El dolor y el miedo, sus emociones homeostáticas que conforman el sentimiento y la motivación, se activan cuando el bienestar de sus cachorros está amenazado. Es como si el círculo dorado del «yo» se ampliara hasta incluir a «mis» indefensos cachorritos (véase figura 3.2).6
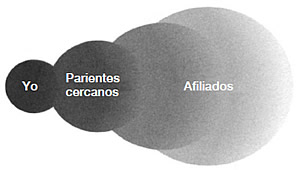
FIGURA 3.2. Esta ilustración muestra las distintas esferas del cuidado. El circuito que permite nuestra propia supervivencia y bienestar en los mamíferos sufre modificaciones hasta abrazar a nuestra prole. En los mamíferos sociales, este abrazo puede incluir a nuestros parientes más cercanos, a miembros de otros grupos e incluso a desconocidos, siguiendo pautas decrecientes de intensidad según el grado de apego.
Si bien es cierto que un lobo o una rata suelen abandonar a sus cachorros cuando se percibe una amenaza tan grande que ella misma debe ponerse a salvo, aunque ello implique no salvar a sus cachorros. Así pues, la ampliación de su ámbito homeostático hasta incluir a los cachorros sigue permitiendo reconocer la distinción entre uno mismo y su querida descendencia. Asimismo, los progenitores humanos que se enfrentan a un enemigo sobrecogedor pueden optar por salvarse a sí mismos, aunque a veces la motivación para salvar a los más jóvenes puede llevar al autosacrificio del progenitor. La conducta humana en estas difíciles circunstancias también dependerá de otros muchos factores, como la naturaleza de la calamidad, el temperamento del individuo, el trasfondo sociocultural y la existencia o no de otros hijos. Son sistemas muy poderosos que interaccionan con el sistema de autopreservación, pero que al mismo tiempo van más allá de él. Ese «ir más allá» no ocurre al azar, sino que se relaciona sistemáticamente con el bienestar de otras personas, especialmente nuestros parientes.7
Los pasos cruciales que nos llevan de cuidar sólo de nosotros mismos a una variedad de tipos de sociabilidad (el cuidado de los demás), que es un rasgo típico de los mamíferos, dependen de los mecanismos neuronales y corporales que «maternalizan» el cerebro de los mamíferos hembra, que a su vez depende de los neuropéptidos como la oxitocina (OXT) y la vasopresina arginina (VPA), junto con otras hormonas. Es casi seguro que estos mecanismos no fueron, en un principio, seleccionados para servir a una serie amplia de finalidades sociales, sino que sólo pretendían asegurar que la hembra contara con los recursos y la motivación necesarios para amamantar, defender y, en términos más generales, dedicarse al bienestar de sus indefensos pequeños hasta que pudieran valerse por sí mismos. Los mamíferos cuyo circuito los preparaba para el cuidado de su descendencia registraban un mayor índice de supervivencia de sus crías que los que se mostraban más negligentes en su cuidado.
Sin embargo, la modificación que nos lleva a cuidar de los demás, básicamente los que forman parte de nuestra descendencia, puede experimentar modificaciones adicionales, tal vez de carácter menor, hasta extenderse al cuidado de individuos que no son de nuestra propia descendencia pero cuyo bienestar es de alguna manera significativo para el bienestar propio y el de nuestra descendencia. Según la especie y su presión de selección, se elije a favor de distintas disposiciones de tipo social, y en ese caso intervienen otros muchos mecanismos cerebrales. De este modo, en una manada de lobos o en una colonia de castores, sólo encontramos una pareja reproductora; en los grupos de babuinos y en los de orcas, todas las hembras fértiles son reproductoras. Los lémures de cola anillada son matrilineales; las hembras son dominantes en relación a los machos, y se emparejan con múltiples machos. En las nutrias de río y los osos pardos, las manadas están compuestas de una hembra y sus cachorros, y la hembra procrea con cualquier macho apto que la corteje adecuadamente. En el caso de los macacos Rhesus, las crías sólo se relacionan con la madre, mientras que los monos tití están más unidos al padre que a la madre. Ésta es sólo una pequeña muestra de la gama de patrones sociales que encontramos en los mamíferos, pero en el trasfondo de todo esto encontraremos probablemente distintos patrones de receptores de oxitocina y otras hormonas y sustancias neuroquímicas.
Los neurocientíficos Porges y Carter plantean la pregunta de por qué la OXT y la VPA deben adaptarse a sus funciones especiales dentro del cerebro de los mamíferos.8 A modo de respuesta, señalan que estos péptidos son sumamente antiguos (tienen al menos setecientos millones de años y anteceden a los mamíferos), y que además están relacionados con la regulación del agua y de las sustancias minerales en los organismos de los animales terrestres en general. Una versión evolutivamente anterior de la oxitocina y la vasopresina —la vasotocina— desempeña una función en la conducta de apareamiento entre anfibios y es importante para los animales que ponen huevos. Mucho antes de la aparición de los mamíferos, la vasotocina formaba parte del juego reproductor. En los mamíferos, la regulación del agua y los minerales se hizo mucho más sofisticada, ya que durante el embarazo, la hembra tenía que desarrollar una placenta y un saco de líquido amniótico lleno de un fluido en el que crecen los bebés, y después del nacimiento la hembra tenía que producir leche para esos bebés.9 Esto nos indica en términos generales por qué la OXT y la VPA se prestan a mutaciones evolutivas dentro del campo de la reproducción de los mamíferos, y por qué los homólogos de estos péptidos son importantes en la sociabilidad de las aves. Tal y como ha observado el biólogo James Hunt, «la sociabilidad, al igual que la multicelularidad, ha aparecido en numerosas ocasiones, en diversas taxonomías y ha alcanzado niveles muy distintos de integración».10 No podemos suponer que todas esas formaciones requieren una OXT y una VPA, y para aquellas que sí las necesitan, el modo en que las emplean puede ser muy dispar.
¿Cómo se consigue, a nivel neural, el apego en los mamíferos? Con el fin de seguir avanzando en esta cuestión, primero debemos explorar con mayor detalle el desarrollo de este concepto de «apego».
VALORES FAMILIARES: PERTENENCIA Y DESEO DE PERTENENCIA
En todas las mamíferas embarazadas, incluidas las mujeres humanas, la placenta del feto libera una variedad de hormonas al torrente sanguíneo de la madre que surte el efecto de «maternalizar» su cerebro.11 Estas hormonas, incluidas la progestina, el estrógeno y la prolactina, actúan principalmente sobre las neuronas de las estructuras subcorticales.12 En los roedores y los gatos, por ejemplo, esta conducta provoca que la hembra embarazada coma más, prepare un nido para su camada y encuentre un lugar considerado seguro para dar a luz. Las hembras humanas también responden a ese impulso del «nido» a medida que se acerca el momento de dar a luz, y (puedo dar fe de ello a título personal) empiezan a limpiar la casa con ahínco y a ultimar los preparativos para la llegada del nuevo bebé. La producción de OXT se regula al alza (es más abundante) durante el embarazo; en el momento del nacimiento, la liberación de OXT participa de la contracción del útero. La OXT también es fundamental en la subida de la leche durante la lactancia. En el cerebro, la liberación de OXT desencadena la plena conducta maternal, que incluye la preocupación por los bebés, amamantarlos y ocuparse de que estén calientes, limpios y seguros. En los seres humanos, la conducta maternal también se activa cuando una mujer adopta a un bebé, y ese apego puede ser tan poderoso como el apego que se siente hacia un bebé al que se haya gestado y alumbrado.13 Es muy probable que la liberación de oxitocina también intervenga en esta conducta. Las tías de las crías de suricatas también responden de este modo. Otros mamíferos, con bebés propios, pueden cuidar de las crías de otras especies, como cuando una perra se deleita en dar de mamar a un cerdito o un gatito.
Es probable que los opiáceos endógenos, es decir, las moléculas parecidas al opio que se generan en nuestro cerebro, también desempeñen un papel crucial en las relaciones maternas, y la hembra que amamanta obtiene la recompensa del placer a partir de los opiáceos que se liberan durante la lactancia. Según mi experiencia, diría que la lactancia es agradable y tranquilizadora, pero no te provoca ninguna clase de «subidón». Las madres de los macacos Rhesus a los cuales se administra naloxona, una sustancia química que bloquea los receptores de opiáceos y, por tanto, también bloquea su efecto, muestran indiferencia hacia sus crías, y tienden a mostrarse negligentes con ellas. Las ovejas a las que se les inyecta naloxona rechazan activamente a sus corderitos. Aunque existen factores sociales que complican la comparación, las adictas humanas a la heroína tienden en su mayoría a mostrarse negligentes con sus hijos o a abandonarlos. Cabe suponer que en las personas que sufren adicción, el efecto modesto de los opiáceos endógenos queda aniquilado por los abrumadores efectos de las cantidades relativamente grandes de heroína,14 aunque los niveles anormales de OXT también pueden tener algo que ver. Las madres que consumen cocaína, por ejemplo, registran niveles más bajos de OXT que los grupos de control de madres no adictas, y además muestran una conducta menos maternal.15 Sin embargo, por lo general, atender a un bebé es una recompensa; sienta bien. En cambio, los niveles de ansiedad se disparan cuando el bebé llora, se separa de la madre o sufre, porque son situaciones que sientan muy mal.
Estas consideraciones nos conducen al tema del dolor, o, en términos más generales, al afecto negativo, que es un aspecto central en el nacimiento de la conducta social de los mamíferos.16 Aunque el dolor puede parecer una reacción muy sencilla cuando se experimenta, cabe añadir que se apoya en una anatomía asombrosamente complicada, que incluye muy distintas especializaciones, componentes, sustancias neuroquímicas, vías y conexiones.17 Aparte de los cambios que requieren oxitocina, el sistema mamífero del afecto negativo —dolor, miedo, pánico, ansiedad— también se modificó. En todos los seres vertebrados, el miedo, la ansiedad y el dolor físico se registran como señales de advertencia en el tallo cerebral y el hipotálamo que incitan a «protegerse». Estos cambios conducen a un repertorio de conductas correctivas a lo largo del circuito de autopreservación. Las modificaciones evolutivas de estos sistemas básicos aseguran que los mamíferos respondan a la amenaza y al ataque a su descendencia como a sí mismos. Las respuestas y los sentimientos implicados en las tácticas de «protegerse a uno mismo» también se hallan en las tácticas de «proteger a los míos».
Un rasgo de firma del cerebro de los mamíferos es la corteza: una superficie compuesta de seis capas uniformes que conforma el borde exterior de los hemisferios cerebrales (véase figura 3.3).18 Este sorprendente invento evolutivo concentra gran parte de la estructura de procesamiento en un espacio restringido, utilizando de forma inteligente una organización microcósmica para sacar el máximo rendimiento a las facultades de procesamiento sin sufrir pérdidas de accesibilidad; es decir, existen conexiones densas a nivel local, conexiones sueltas con regiones lejanas, pero gracias a las excelentes conexiones con los vecinos, cualquier zona puede llegar a otras sólo con dar unos pocos pasos. En nuestra historia, la modificación de un interés particular tiene que ver con la elaboración cortical de la representación del dolor, y en concreto, del dolor que se produce cuando nos separamos de nuestros seres queridos o cuando sufren una amenaza.
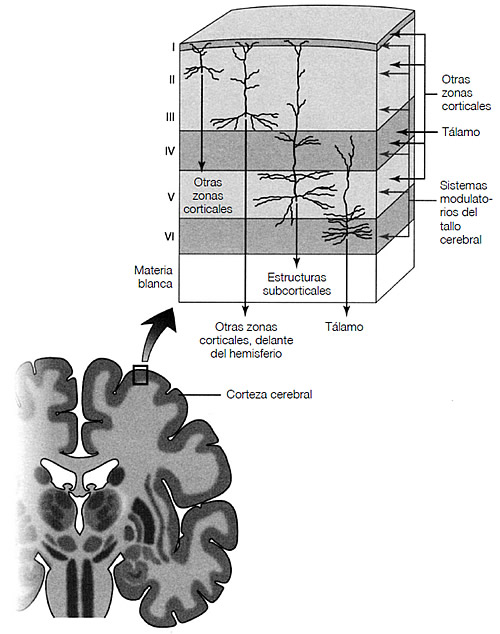
FIGURA 3.3. Vista del cerebro en sección coronal (un corte de una oreja a otra). Los bordes grises de la superficie exterior son la corteza (manto cortical). La diferencia de color entre la materia blanca y la materia gris depende de la presencia de la mielina, que está compuesta de células ricas en grasa que se envuelven alrededor de los axones de las neuronas, proporcionando un tipo de aislamiento que facilita una transmisión más rápida de señales. La materia gris carece de mielina. La sección representa la organización laminar de la corteza y su arquitectura altamente regular. No puede apreciarse la densidad de las neuronas; existen unas cien mil neuronas en un milímetro cúbico de tejido cortical, con aproximadamente mil millones de conexiones sinápticas entre ellas. Adaptación de A. D. Craig, «Pain Mechanisms: Labeled Lines versus Convergence in Central Processing», Annual Review of Neuroscience, nº 26, 2003, págs. 1-30, y E. G. Jones, «Laminar Distribution of Cortical Efferent Cells», en A. Peters y E.G. Jones (comp.), Cellular Components of the Cerebral Cortex, Nueva York, Plenum, 1984, vol. 1, págs. 521-553.
Tal y como el neurocientífico A. D. (Bud) Craig reconoce tímidamente, el dolor es un enigma. Aun así, los principales descubrimientos de los últimos quince años, muchos de ellos realizados por Craig y sus colaboradores, han corregido algunas ideas equivocadas que se han ido arrastrando desde hace tiempo, y ha convertido el dolor y todas sus rarezas en un misterio menos impenetrable.19 Al clasificar el dolor como una emoción homeostática, en vez de como una sensación como la presión, Craig pretende hacer hincapié en el amplio conjunto de mecanismos de autopreservación. Esto supone un contraste con la función de la visión y la audición, que principalmente se dedican a representar el mundo externo pero no implican de modo automático una «sensación» motivacional del mismo modo que lo hace, por ejemplo, una quemadura. Desde luego, el contraste es una cuestión de grado, puesto que en última instancia la visión y la audición también sirven a la autopreservación —a veces sólo un poco menos directamente—. Al igual que con la sed o el frío, las señales del dolor que proceden de las tripas del cuerpo, los músculos o las articulaciones activan la necesidad de corrección. Como parte de los procesos dinámicos para mantener la homeostasis, el aspecto sensorial del dolor puede variar incluso cuando el estímulo permanece inalterable; por ejemplo, los soldados que reciben un balazo en el fragor de la batalla pueden no sentir dolor hasta que se sienten a salvo en el hospital, del mismo modo que pueden no sentir sed o hambre.
Tal y como aparece reflejado en la figura 3.4, el sistema central del dolor en los seres humanos nace de una región de la médula espinal llamada lámina I, en la que las señales sobre el dolor (nocicepción) se reciben de los tejidos corporales y de los órganos. La vía neural transporta estas señales hasta la médula espinal, en el sistema cerebral, donde se produce un diálogo entre regiones que regulan las respuestas homeostáticas, y después continúa hasta las regiones específicas del tálamo. Este sistema, conocido como tracto espinotalámico, permite la localización muy precisa de señales nociceptivas, así como distintas clases de sentimientos dolorosos —la diferencia entre un dolor punzante y una sensación de quemazón o la sensación de pérdida—. A nivel cortical, hay dos lugares que desempeñan un papel crucial en el procesamiento del dolor: la ínsula (arremolinada debajo del lóbulo frontal y que puede pasar desapercibida) parece un elemento esencial en el procesamiento de las sensaciones desagradables que nos aportan las experiencias dolorosas —los aspectos cualitativamente negativos— mientras que la corteza cingulada anterior (CCA), a la cual está conectada, domina el aspecto motivacional (el de «hacer» algo) del dolor. La ínsula, tal y como aparece en la figura 3.5, integra la sucesión de señales corporales para que envíen un informe completo sobre el estado del organismo.
Primero, las señales llegan a la parte posterior de la ínsula, y luego parece que se reprocesan a través de una serie de pasos que van desde la parte de atrás al área frontal de la ínsula, y probablemente asciendan en complejidad e integración según lo que se intenta representar. La ínsula parece representar el «estado de uno mismo» y «el estado de lo mío», ya que integra las señales procedentes de todo el cuerpo y del cerebro. Cuando se registra una señal de que «algo va mal», como un frío paralizante o un ataque inminente, el sistema responde con señales de incomodidad o dolor, motivando así una acción de respuesta. Siendo fieles a su neuroanatomía, los pacientes con demencia frontotemporal, que implica la destrucción de las neuronas situadas en la ínsula, muestran una pérdida pronunciada de respuestas empáticas así como una disminución de la experiencia del dolor.
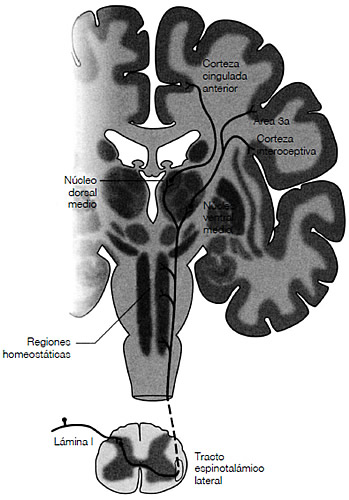
FIGURA 3.4. La ilustración representa la vía dominante que sigue el dolor en el cerebro humano y la médula espinal tal y como se vería en una sección coronal. La corteza y otras estructuras de materia gris (principalmente los cuerpos de neuronas) aparecen sombreadas con un gris más oscuro; la materia blanca (principalmente, axones mielinizados de neuronas), con un gris claro. Obsérvese que el tracto espinotalámico lateral realiza conexiones en el tallo cerebral con la región que regula la homeostasis, y luego continúa estableciendo conexiones sinápticas en dos núcleos distintos (regiones de materia gris) en el tálamo. Un núcleo talámico se proyecta hacia la ínsula anterior (la corteza interoceptiva), que contiene una representación del estado fisiológico del cuerpo, y hacia la corteza somatosensorial (área 3a); el otro núcleo envía neuronas al CCA (corteza cingulada anterior). Extraído de A. D. Craig, «Pain Mechanisms: Labeled Lines versus Convergence in Central Processing», Annual Review of Neuroscience, nº 26, 2003, págs. 1-30. (Reproducido con el permiso del autor.)
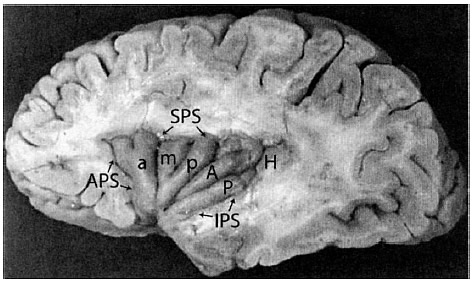
FIGURA 3.5. Una fotografía anatómica que muestra la ínsula del hemisferio izquierdo del cerebro. La ínsula ha quedado expuesta al diseccionar partes del lóbulo frontal, el lóbulo temporal y el lóbulo parietal. La ínsula también puede quedar expuesta sin ser diseccionada separando el lóbulo frontal del temporal. a, m, p: giro anterior, medio y posterior de la ínsula anterior; A, P: giro anterior y posterior de la ínsula posterior; APS: surco periinsular anterior; SPS: surco periinsular superior; IPS: surco periinsular inferior; H: giro Heschel. (Un «giro» o «circunvolución» es un montículo; un «surco» es una hondonada; esta geografía viene dada por el efecto de plegado a medida que el cerebro crece y se amolda a la estructura del cráneo. Los puntos de referencia son aproximados y varían sensiblemente de una persona a otra.) Extraído de Thomas P. Naidich y otros, «The Insula: Anatomic Study and MR Imaging Display at 1.5 T», American Journal of Neuroradiology, nº 25, 2004, pág. 226. (Reproducido con el permiso del autor.)
La región situada en la parte superior de la jerarquía de procesamiento —la ínsula anterior— parece ser un rasgo único de los primates, y está más desarrollada en los humanos que en otros primates.20 No se sabe con exactitud lo que estas diferencias significan en cuanto a la disparidad de capacidades, pero pueden incidir en la complejidad de representación del «estado del yo» y el «estado de lo mío».21
Puesto que los seres humanos tienen cerebros sociales, nuestro sistema de dolor más generalizado nos hace sentir mal no sólo cuando nuestro bienestar se ve amenazado, sino cuando el bienestar de nuestros seres queridos también lo está. Las crías de mamífero están asustadas cuando se separan de quienes sienten apego, y emiten llamadas de socorro. Se trata de un rasgo positivo, puesto que no pueden alimentarse por sí mismos ni defenderse, y necesitan a su madre o a su padre.22 Además, las madres mamífero, y, en algunas especies, los padres, sienten ansiedad y malestar cuando oyen a sus crías emitir llamadas de ayuda. También es un rasgo positivo por la misma razón: sus crías los necesitan. Tanto la ínsula como la CCA responden al dolor físico, pero también responden al dolor social motivado por la separación, la exclusión o la desaprobación, así como al dolor que resulta de errores y predicciones incorrectas.23
Cuando una madre mamífero logra poner a salvo a su bebé y satisfacer sus necesidades, se liberan opiáceos endógenos y OXT, tanto en el cerebro del bebé satisfecho como en el de la madre satisfecha. Estar juntos les sienta bien. Los humanos conocemos este sentimiento aunque no sepamos nada acerca de la oxitocina ni de los opiáceos endógenos.
Los sentimientos de remordimiento, culpa y vergüenza son típicos de la mayoría de seres humanos (no todos) después de dañar a alguien. Los psicópatas, aunque conozcan la importancia social de expresar remordimientos durante un juicio, en realidad no sienten ninguno, aunque hayan causado terror, mutilaciones y muerte. Los psicópatas son personas que pueden ser astutos y encantadores en un determinado entorno social pero que carecen de conciencia, y no tienden a crear vínculos afectivos.24 Los estudios que existen sobre los psicópatas (las palabras «psicópata» y «sociópata» suelen emplearse como sinónimos) nos han permitido sintetizar una serie de criterios muy precisos para su diagnóstico.25 Los individuos a quienes se les diagnostica una psicopatía suelen tener seis o más delitos graves en su historial delictivo, son reacios a formar relaciones a largo plazo, son manipuladores, embaucadores, y no tienden a experimentar sentimientos profundos, sean positivos o negativos. Ted Bundy, que confesó haber cometido treinta asesinatos entre 1974 y 1978 y que se dedicó a la tortura y a la necrofilia, fue el clásico ejemplo de psicopatía. No sentía ningún tipo de remordimiento o culpa por sus acciones, pero en cambio era un tipo atractivo y encantador. Por el contrario, Charles Manson, el líder de una secta acusado de conspiración para cometer los asesinatos de Tate y LaBianca en Los Ángeles en 1969, sufría un espejismo, y se veía a sí mismo como el líder de una revolución que beneficiaría a la humanidad.
¿Son distintos los cerebros de los psicópatas? Por lo visto, sí. Los datos de los que disponemos indican diferencias importantes entre los cerebros de los psicópatas y los de los sujetos de control sanos en las zonas que regulan las emociones, los impulsos y las respuestas sociales. En concreto, las regiones paralímbicas del cerebro son distintas en los psicópatas tanto a nivel anatómico (son de un tamaño más reducido) como funcional (registran niveles más bajos de actividad en el aprendizaje emocional y en las tareas de toma de decisiones).26 Las zonas paralímbicas incluyen las áreas que uno esperaría que estuviesen implicadas: estructuras subcorticales que regulan las respuestas emocionales, como la amígdala y el septo, estructuras relacionadas con la memoria (las zonas del hipocampo), y las zonas de la corteza que se sabe que participan de las interacciones sociales, incluidos los sentimientos de dolor y placer social (la ínsula, la CCA, la corteza orbital frontal y el lóbulo temporal lateral; véase figura 3.6).27
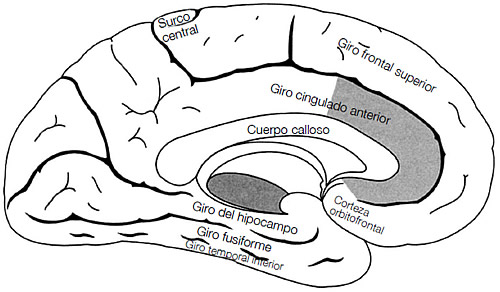
FIGURA 3.6. Dibujo del cerebro humano en el que se muestra la ubicación de la corteza cingulada anterior, la corteza orbitofrontal (llamada así porque está situada encima de las órbitas de los ojos), el giro del hipocampo, el giro frontal superior, el giro temporal inferior, el giro fusiforme y el cuerpo calloso (la principal vía de conexión entre los dos hemisferios). Basado en Wikimedia Commons (<http://en.wikipedia.org/wiki/indez.php?title=Special%3ASearch& search=anterior+cingulate>).
Los estudios en gemelos y en miembros de una misma familia indican que la psicopatía es hereditaria en un rango aproximado del 70%; las condiciones de abuso o desatención durante la infancia pueden agravar la psicopatía de quienes están predispuestos genéticamente a ella.28 Puesto que una parte muy importante de la población carcelaria (quizá un 30 o un 40%) registra patrones elevados en los criterios para diagnosticar la psicopatía, y debido a que los psicópatas pueden ser embaucadores y destructivos, nos encontramos ante un trastorno social de gran calado. La psicopatía también nos recuerda la importancia del afecto negativo en nuestras relaciones sociales —su papel crucial en el aprendizaje de una conducta social adecuada, en acallar las acciones antisociales y en desarrollar una conciencia.
Si no se siente dolor social, entonces causar una mutilación que acabe con una vida tiene la misma importancia que una broma de colegial.29
Puesto que los cerebros humanos poseen amplias regiones prefrontales y límbicas, no necesitamos responder con rigidez a situaciones desagradables.30 Podemos considerar otras opciones que nos permitan evitar el dolor en un futuro, o podemos soportar un dolor inmediato para obtener un beneficio posterior. Podemos trazar planes a largo plazo y pensar en sus posibles consciencias. El modo en que efectuamos todas estas operaciones es un tema de constante investigación, aunque aún no alcancemos a comprenderlo del todo.31 Lo que sí sabemos es que aplazar la gratificación suele implicar un grado de conciencia así como el estímulo de la imaginación al pensar en los efectos que tienen nuestras acciones en el futuro, pero también pueden influir hábitos arraigados que estén instalados en nuestro sistema de recompensas.32 Sin embargo, esta evaluación no debería considerarse como una cuestión «estrictamente cognitiva», en el sentido de no estar influenciada por las emociones, como lo sería el cálculo de una suma de las cifras 29 y 57. De hecho, la valoración de acontecimientos futuros imaginados, un proceso de restricción y satisfacción, está moldeada por señales del circuito de valencias que constituye la esencia misma del ser, de sentirse bien y de desenvolverse socialmente. La «fría» razón no está desprovista de emociones. Se diría que queda equilibrada por las emociones de actitudes como la prudencia, la vigilancia y la cautela.
Hay otra modificación evolutiva en el cerebro de los mamíferos, que implica al nervio vago y al tallo cerebral (véase figura 3.7). El nervio vago es una clase de conducto que permite enviar y recibir señales detalladas de todos los aspectos del cuerpo —interno, muscular, piel y huesos—. Los mamíferos desarrollaron una nueva rama del nervio vago que modificó de un modo profundo una respuesta especializada de conducta ante el peligro, a saber: la paralización. Quedar paralizado puede confundir a un depredador, ya que depende del movimiento para saber dónde se encuentra exactamente la presa. Una lagartija puede quedarse quieta si se la asusta, por ejemplo. El neurocientífico Stephen Porges33 afirma que las modificaciones en los mamíferos del circuito de «congelación» permite una nueva conducta que mantiene la parálisis pero expulsa el miedo asociado a ella, permitiendo de este modo la inmovilidad sin temor.
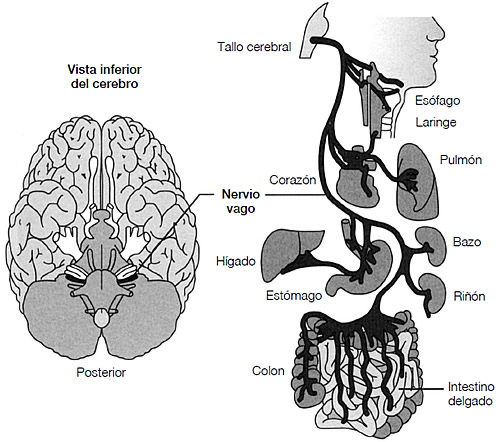
FIGURA 3.7. Las vías del nervio vago. Izquierda: diagrama esquemático que muestra la ubicación del nervio vago (el décimo nervio craneal) cuando entra en el tallo cerebral, visto desde la parte inferior del cerebro. Derecha: diagrama esquemático que muestra la gama excepcionalmente amplia de inervación por el nervio vago. Se cree que las «intuiciones» dependen de las señales del nervio vago. Copyright Bloomsbury Educational Ltd., <www. clinicalexams.co.uk/cranial-nerves-system.asp>. (Adaptado con permiso.)
¿Por qué este factor es importante? Porque las madres mamífero necesitan la capacidad de estar inmóviles sin sentir temor mientras permanecen vigilantes. Desde la perspectiva de los reptiles, ésta es una extraña combinación de estados, pero para amamantar a las crías durante varias horas al día, una madre mamífero necesita quedarse muy quieta para que las crías puedan alimentarse sólo de su alimento.
Su cuerpo no puede estar en «modo de apagado», sino preparado para responder a amenazas y a intrusos. Cuando las madres mamífero están tranquilas, el cuerpo y el cerebro no deben responder como si estuvieran paralizadas con miedo, porque paralizarse con temor movilizaría el sistema simpático, amortiguando los efectos de la oxitocina, e interrumpiendo así el flujo de la leche. La madre lactante debe estar quieta, calmada y relajada, pero también preparada por si surge un peligro. La madre mamífero también necesita estar tranquila para la cópula (pensemos en las vaquillas que básicamente se quedan quietas mientras el toro deposita enérgicamente su esperma) y para el parto (correr sería peligroso para la madre o la cría). De este modo el nervio vago, que a menudo se tiene por otro aburrido y viejo nervio craneal que se debe memorizar para el examen de anatomía, es, en el caso de los mamíferos, una parte muy especial de nuestra naturaleza social.
Para las ratas, los ratones y otros muchos mamíferos con una corteza prefrontal (CPF) pequeña en comparación con el tamaño del cuerpo, el tallo cerebral y otras estructuras subcorticales son los elementos principales en la integración de señales y en la toma de decisiones, como si se debe huir ahora y comer después, si se debe luchar ahora o dentro de unos días, o si se debe incurrir en una experiencia dolorosa para defender a las crías de un depredador. En los primates, como los monos, los chimpancés y los humanos, la amplia corteza prefrontal implica que las estructuras subcorticales participan del proceso de toma de decisiones pero no suelen dominar en él, aunque puede ser así en episodios de pánico.34 De ahí que, en los primates, exista una relación más flexible y más compleja entre los estímulos y la conducta. Barry Keverne ha descrito esta holgura como una especie de liberación de los patrones fijos de acción que vemos en los mamíferos de cerebro pequeño.35
La incursión en la compleja espesura del dolor, el miedo, el placer y el sistema de recompensas tiene una contrapartida más. La predicción, tal y como señala el neurocientífico Rodolfo Llinàs, es la función cerebral más importante y más generalizada de todas.36 Esto es así porque a la hora de guiar la conducta, las operaciones predictivas están al servicio de la supervivencia y el bienestar. Cuanto mejor sea la predicción, más probabilidades hay de que el individuo sobreviva a un depredador, encuentre buena comida y evite los peligros. Al igual que en la magia del interés compuesto, las capacidades predictivas ganan exponencialmente en capacidad y abstracción gracias a la expansión de las redes neurales entre la entrada sensorial y la salida motora. Los primeros mamíferos podían utilizar su neocorteza para anticiparse con mayor efectividad a una circunstancia potencialmente peligrosa. Los mamíferos de gran cerebro pueden ser incluso más inteligentes en sus predicciones y en su modo de actuar.37 Para los mamíferos sociales, anticiparse a lo que harán los demás es sumamente valioso: ¿el otro compartirá, morderá, pegará, se emparejará o qué hará?
Saber anticiparse a un problema social suele implicar una carga emocional, y motiva la acción preventiva de un modo adecuado a las circunstancias. Además, el bebé aprende a anticiparse a la conducta de su madre y a la de sus compañeros de camada, llegando incluso a predecir las acciones de ciertos movimientos depredadores —juego, heridas, etcétera—. Según proponen Don Tucker y sus colaboradores, esta dinámica marca el modesto inicio de una representación interna de los objetivos de otros individuos,38 una representación que es más abstracta que una predicción de movimiento pero que depende de haber aprendido una serie de asociaciones mediadas por las neuronas neocorticales.39 Desde hace muy poco tiempo, los experimentos en conducta animal se han centrado en descubrir si los animales no humanos tienen un modelo mental de los objetivos y los puntos de vista de terceros, y algunos de los resultados son asombrosos. Según ha demostrado la etóloga Nicola Clayton, un arrendajo entiende lo que otros arrendajos pueden ver, y es capaz de adaptar su estrategia de caza como consecuencia de ello.40 Si un arrendajo que esté en posición predominante puede ver el alijo de bellotas de otro arrendajo, el propietario de esos alimentos mueve el alijo. Pero no lo hará si el arrendajo que observa no es predominante dentro del grupo.
Los chimpancés han demostrado ser totalmente capaces de ajustar su conducta de un modo muy parecido al de los arrendajos. Pensemos en un chimpancé madre que anticipa que su cría provocará una reacción hostil en el macho alfa si esa cría se atreviera a coger parte de la comida destinada a los adultos. La madre siente cierto dolor anticipatorio al detectar las intenciones de la cría, y la ahuyenta antes de que empiecen los problemas. En los seres humanos, estas humildes pero útiles herramientas predictivas conducen a un amplio esquema de los estados mentales de los demás; a saber, una «teoría de la mente» repleta de representaciones abstractas como son los «objetivos» y las «creencias». (Me referiré a la neurobiología de la «teoría de la mente» en los capítulos 4 y 6 de este libro.)
La creciente comprensión neurobiológica del apego en los mamíferos encaja con la observación de que el sistema nervioso conserva un gran parecido entre especies. Las neuronas son muy parecidas y presentan un funcionamiento muy similar en los humanos, los ratones y las babosas; la paleta de sustancias neuroquímicas que incide en las neuronas y los múscu los es sustancialmente la misma en todos los vertebrados e invertebrados; los patrones básicos del desarrollo del cuerpo y del cerebro también son parecidos entre los vertebrados y los invertebrados. Lo más curioso es que las modificaciones más modestas en las estructuras neurales existentes —la expansión de una zona auditiva o la ampliación de una región que repre sente el tacto de los dedos, por ejemplo— pueden generar resultados dis tintos, como una mayor capacidad para discernir patrones de sonidos o de tacto.41 En este sentido, el neurobiólogo Jaak Panksepp sugiere que el do lor que sienten los mamíferos ante una separación social puede deberse a una modificación de un antiguo instinto de «preferencia por un lugar» —con su correspondiente ansiedad ante un espacio desconocido— que suele apreciarse en los animales no mamíferos.42 La familiaridad es agrada ble porque permite una mayor predictibilidad, y ello conlleva una reduc ción de la ansiedad. El placer de estar en compañía de otros miembros de un mismo grupo explota el circuito de sentirse cómodo en unas condicio nes conocidas y seguras, es decir, en un estado de «reposo y digestión». Los cambios evolutivos de los mamíferos, en virtud de los cuales se siente do lor ante la separación de nuestros seres queridos, son tal vez leves modifi caciones desde el punto de vista del circuito cerebral, pero generan algo bastante distinto en un nivel más amplio: procurar cariño a los demás.
La evolución biológica no logra adaptaciones diseñando un nuevo mecanismo desde el principio, sino que modifica poco a poco lo que ya existe. Las emociones sociales, los valores y la conducta no son el resulta do de un plan de ingeniería completamente nuevo, sino más bien una adaptación de patrones existentes y mecanismos que están íntimamente relacionados con el circuito de autopreservación para la lucha, la paraliza ción y la huida, por un lado, y para el reposo y la digestión, por otro. El dolor de la exclusión, la separación y la desaprobación, por ejemplo, no requiere un nuevo sistema, sino que explota, amplía y modifica lo que ya existe para registrar el dolor físico y las emociones homeostáticas en los mamíferos. En el próximo apartado analizaremos cómo el sistema nervioso de algunos mamíferos amplió sus patrones de apego más allá del círculo restringido de su descendencia.
APEGO A LAS PAREJAS
Aunque a veces damos por sentado que se trata de un patrón único que caracteriza a los seres humanos, lo cierto es que el apego a largo plazo a una pareja se halla en un 3% de todos los mamíferos, incluidos los castores, los marmosetas, los macacos tití, los gibones, el ratón de patas blancas, los ratones de campo y el ratón de los pinos.43 Sin embargo, la mayoría de mamíferos, aunque sean sociales, son promiscuos o estacionales en sus pautas de emparejamiento. Una proporción mucho más elevada de aves (cerca del 90%) sienten una fuerte preferencia por su pareja y mantienen relaciones a largo plazo.44 Nuestros parientes vivos más cercanos (los chimpancés y los bonobos) no establecen relaciones de pareja a largo plazo, y lo mismo ocurre en la mayoría de roedores y monos.
El apego a largo plazo a una pareja es una forma altamente significativa de sociabilidad: amamos al otro, queremos emparejarnos con él, estar juntos, verle prosperar y cuidarlo. Nos entristecemos durante la separación o cuando nuestra pareja está herida o amenazada. Cuando una pareja fallece, el miembro superviviente se deprime y a veces le cuesta mucho recuperarse de esa tristeza.45 Sin embargo, el apego a una pareja no implica exclusividad sexual —tal y como revelan los estudios genéticos en roedores y humanos—,46 lo cual podría estar relacionado con la diversidad genética, tal y como sugieren los estudios sobre el topo común (que tiene el evocador nombre científico de Cryptomys hottentotus hottentotus).47 Pero, actualmente, la cuestión que se baraja es la siguiente: cuando detectamos una intensa preferencia por una pareja en los ratones de campo, por ejemplo, ¿qué información procesa el cerebro para que un ratón de campo sienta inclinación por una misma pareja de por vida, pero no ocurra lo mismo, en cambio, en un ratón de montaña?
Los ratones son roedores cuyo aspecto es muy parecido al de los ratones gorditos con colas cortas. Los ratones de campo y de montaña, por mucho que se parezcan físicamente, son muy distintos en cuanto a sus hábitos sociales: los ratones de campo se emparejan de por vida; los de montaña no muestran ninguna preferencia por sus parejas. Los ratones macho de campo resguardan a la hembra y la protegen de posibles intrusos, y los machos comparten el cuidado de las crías, lamiéndolas, moviéndolas y defendiéndolas. En el caso de los ratones de montaña, sólo las hembras cuidan de sus crías, y lo hacen por un período más breve que en los ratones de campo. Los niveles generales de sociabilidad también son distintos. Si los colocamos al azar en una sala amplia, los ratones de campo tenderán a permanecer juntos; en cambio, los de montaña se conforman con estar a solas. Las parejas macho-hembra de ratones de campo proporcionan la base para crear grupos familiares más amplios, en los que los parientes ayudan en el cuidado de los más jóvenes. No ocurre así en los ratones de montaña.
Los ratones de montaña y los de campo son muy parecidos en la estructura general de sus cerebros, pero si los comparamos a nivel microestructural descubriremos diferencias en su neurobiología que explicarían estas grandes diferencias en sus hábitos de sociabilidad.
En la década de 1970, Sue Carter, una neuroendocrinóloga de la Universidad de Illinois, había estado estudiando los efectos de las hormonas en el cerebro y la conducta cuando observó que los ratones de campo muestran preferencias por una pareja y que, además, el vínculo afectivo se forma en el primer emparejamiento. Mientras se preguntaba acerca de este sorprendente fenómeno, llegó a pensar que las hormonas sexuales —probablemente el estrógeno— tenían la clave para explicar las inusuales pautas de apego de los ratones. Aunque fue una suposición acertada, sus experimentos no corroboraron la hipótesis del estrógeno. Buscó la respuesta en otra parte, y reflexionó acerca del extraordinario trabajo neuroendocrinológico del biólogo Barry Keverne realizado en ovejas.48 El laboratorio de Keverne había demostrado que una inyección del neuropéptido oxitocina en el cerebro de una oveja sexualmente inexperta podía suscitar una conducta maternal plena, incluido un vínculo entre oveja y cordero. Como bien sabrá cualquier granjero, conseguir que una oveja se una afectivamente a un corderito huérfano es muy difícil, incluso en el caso de las ovejas que acaban de dar a luz a un cordero muerto y estarían dispuestas a cuidar de otro. El resultado era, por tanto, sorprendente. Si el apego entre madre y crías está regido por la oxitocina, ¿podríamos afirmar que los vínculos afectivos con la pareja son una extensión de esta misma pauta? El instinto de Carter resultó ser de gran utilidad, y puso en marcha una serie de investigaciones al respecto. Mientras leía acerca de estos estudios, empecé a sospechar que éste era el vínculo asociado con el tipo de cuidados que normalmente relacionamos con la conducta moral humana.49 Quizá Hume lo aceptaría como el germen del «sentimiento moral».
LOS MECANISMOS DEL APEGO A LA PAREJA
Tal y como mencioné anteriormente, la oxitocina es un péptido muy sencillo y muy antiguo; se trata de una cadena compuesta de nueve eslabones de aminoácidos (la hemoglobina, en cambio, es una cadena altamente compleja, con más de quinientos eslabones de aminoácidos). La molécula de la OXT tiene un pariente, la vasopresina arginina (VPA), y ambas parecen haberse desarrollado a partir de un antepasado común, del que sólo diferían en dos aminoácidos. Al igual que la OXT, la VPA se halla en el cerebro y en el organismo, donde desempeña un papel fundamental en el mantenimiento de la presión sanguínea y el equilibrio del agua.
La OXT y la VPA se liberan naturalmente en el hipotálamo, y de ahí se esparcen ampliamente a otras áreas subcorticales, como las que están implicadas en las dinámicas de recompensa (incluido el núcleo accumbens), en la regulación de la conducta sexual (el septo) y en la regulación de los hábitos de parentesco. La OXT es más abundante en las hembras que en los machos. La VPA se libera a partir de otras áreas subcorticales, incluyendo la amígdala medial, el septo lateral y el núcleo periventricular, y es más abundante en machos que en hembras. Los niveles de VPA aumentan cuando el macho entra en la pubertad; se libera durante la etapa de excitación sexual y decae poco antes de la eyaculación. En las ratas macho, la OXT favorece la función eréctil, llegando a un nivel máximo durante el orgasmo y descendiendo a niveles mínimos al cabo de treinta minutos después de dicho orgasmo.50 El factor de liberación de corticotropina (FLC) también es una pieza muy importante de la vida social de los mamíferos. Se relaciona con el estrés, y por tanto con la ansiedad y sus incomodidades. Cuando los animales se sienten seguros y cómodos, sus niveles de OXT suben y sus niveles de FLC descienden del nivel de «lucha o huida». Por consiguiente, se reduce la ansiedad. Sin embargo, cabe destacar que niveles moderados de FLC incrementan los lazos afectivos en los ratones macho de campo.51
Con el fin de hacer sentir sus efectos, la OXT y la VPA se vinculan a receptores específicos de proteínas en la superficie de las neuronas, y de ahí que el papel de la OXT en la conducta dependa de su abundancia relativa, pero también de la densidad de los receptores de las neuronas en una zona particular del cerebro. Por ejemplo, las ratas hembra que lamen y cuidan más de sus crías presentan una mayor densidad de receptores de OXT que otras hembras. La OXT sólo tiene una clase de receptor, pero la VPA tiene dos receptores distintos en el sistema nervioso. Vincularse a uno significa desempeñar una función determinada en los lazos afectivos del emparejamiento y la conducta de los progenitores respecto a su prole, mientras que vincularse al otro se relaciona con la ansiedad y la agresión, especialmente en cuestiones de protección de la pareja.52
En las investigaciones llevadas a cabo hasta la fecha, el principal contraste neurobiológico entre los ratones macho de montaña y los ratones macho de campo es que estos últimos tienen una densidad muy superior de receptores de VPA y OXT en dos regiones subcorticales muy específicas del cerebro: el pálido ventral y el núcleo accumbens (ambos forman parte del sistema de recompensas y castigo; véase figura 3.8).53 Aunque todos los mamíferos tienen OXT y VPA en el sistema nervioso central y todos tienen receptores para ambos, los experimentos han demostrado que es la densidad del receptor en estas dos regiones concretas y altamente interconectadas lo que determina una diferencia clave en la conducta social. Si a modo de experimento bloqueamos los receptores de manera que la OXT y la VPA no puedan vincularse, entonces los ratones tratados no establecerán vínculos afectivos después del primer emparejamiento, y no mostrarán la conducta social típica de los ratones de campo. Como la neurobiología es lo que es, no debería sorprendernos que otros factores adicionales, como las variaciones en los circuitos neurales y en los niveles de otras hormonas, tuviesen un papel destacado en el hecho de que los individuos de una especie formen lazos afectivos a largo plazo o no. Por eso es mejor considerar los datos sobre las elevadas densidades de receptores que se correlacionan con las relaciones afectivas a largo plazo como el inicio de la historia, no como su final.
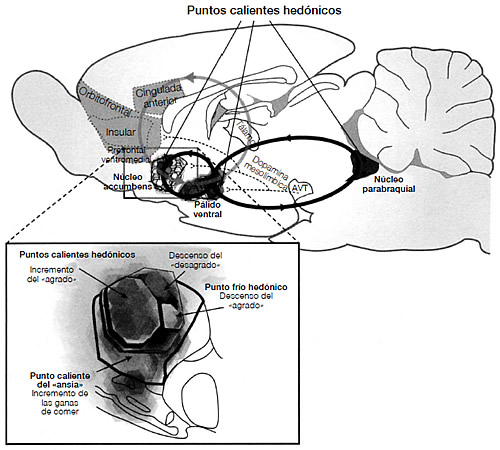
FIGURA 3.8. Dibujo del cerebro de una rata en el que se muestra el circuito principal del sistema de recompensas. Tres estructuras subcorticales cruciales conforman el núcleo accumbens, el pálido ventral y el núcleo parabraquial. Las principales estructuras corticales relacionadas con los puntos calientes hedónicos son la cingulada anterior, la corteza orbitofrontal, la ínsula y la corteza frontal ventromedial. El AVT (área ventral tegmental) contiene neuronas que liberan dopamina, y estas neuronas se proyectan en el pálido ventral, el núcleo accumbens y la corteza orbitofrontal, y son importantes en el aprendizaje del sistema de recompensas. Todas las estructuras y vías también existen en el cerebro humano. Extraído de Kent, C. Berridge y Morten Kringelbach, «Affective Neuroscience of Pleasure: Reward in Humans and Animals», Psychopharmacology, nº 199, 2008, págs. 457-480. (Reproducido con el permiso del autor.)
¿Cuál es exactamente el efecto que se produce en las neuronas cuando estos dos péptidos, la OXT y la VPA, se vinculan a sus respectivos receptores? Ya se están llevando a cabo investigaciones en este sentido, aunque aún no tenemos respuestas concluyentes. Además, las respuestas van a ser sin duda alguna complejas, incluso en el caso de los ratones, puesto que las neuronas que inciden en estos procesos forman parte de un sistema más amplio, lo cual significa que lo que ocurra en otras partes (en el ámbito de la percepción, la memoria, etcétera) también tendrá un impacto. La OXT se libera durante las interacciones sociales positivas, y se ha demostrado que inhibe las conductas defensivas, como la lucha, la huida y la parálisis. Al parecer, lo hace mediante la interacción con el eje hipotalámico-pituitario-adrenal con el fin de inhibir la actividad en la amígdala, una estructura evolutivamente antigua que, entre sus distintas funciones, tiene la de regular las respuestas al miedo. La liberación de OXT también tiende a regular a la baja (a amortiguar) las respuestas automáticas de lucha y huida en el tallo cerebral, y en general reduce la reactividad del sistema nervioso ante los factores que provocan estrés. En consecuencia, sus efectos dependen del contexto en el que se encuentren. La OXT administrada a las ratas macho incrementa la agresión hacia un intruso, pero hace descender la agresión hacia las crías.
¿Se extiende el perfil de densidad de receptores que vemos en los ratones de campo a otras especies monógamas? Al parecer la respuesta es positiva en los monos marmosetas,54 los macacos tití55 y los ratones de patas blancas (Peromyscus californicus).
En cambio, las especies promiscuas como el mono Rhesus y la variedad Peromyscus leucopus del ratón de patas blancas tienen un perfil de receptor de OXT y de VPA parecido al del ratón de montaña (que es promiscuo). Aún no se han determinado datos comparables acerca de la densidad de receptores en la anatomía humana, puesto que los métodos de inyección para etiquetar los receptores no pueden llevarse a cabo en humanos vivos, y no son eficaces cuando se aplican a cerebros de personas fallecidas. Sin embargo, debido a que los mecanismos y las estructuras se conservan bastante bien entre distintas especies, es razonable suponer que los humanos que forman relaciones estables a largo plazo tendrán densidades de receptores más parecidas a las de los ratones de campo, los marmosetas y los gibones que a las de los monos de montaña y los chimpancés.
En su investigación de las relaciones existentes entre los genes, el cerebro y la conducta en humanos, Heike Tost indicó recientemente que una variante particular del gen receptor de OXT (OXTR) se corresponde con algunos tipos de variables en la sociabilidad estudiada en humanos, incluidos algunos tipos de impedimento social.56 El alelo (el gen OXTR se conoce como rs53576 y el alelo como rs53576A) establece una correlación con diferencias anatómicas concretas (relacionadas con los sujetos normales de control): descensos en el tamaño de la materia gris en el hipotálamo; mayor conectividad entre el hipotálamo y la amígdala y entre el hipotálamo y la corteza anterior cingulada; y, sólo en los machos, un incremento en el volumen de la materia gris de la amígdala.57 En las pruebas para determinar si ese factor incidía en la actividad cerebral durante una tarea que supusiera una inversión emocional, los investigadores registraron un descenso en los niveles de actividad de la amígdala.
En cuanto a la conducta, utilizando escalas establecidas de autovaloración,58 el alelo estaba relacionado con una menor sociabilidad (deseo de pertenencia, empatía hacia los demás, paternidad y maternidad responsables, capacidad de crear vínculos afectivos a largo plazo, etcétera). Todavía no existe ninguna técnica para determinar directamente la densidad y la distribución de los receptores en sujetos vivos, de ahí que las investigaciones se centren en estructuras neurales que se sabe que aíslan la OXT (por ejemplo, el hipotálamo) o que están altamente conectadas con áreas que tienen receptores de OXT (por ejemplo, la amígdala). Para explicar las variaciones en el temperamento social, Tost y otros señalan que, en los sujetos que tienen el alelo rs53576A, la estructura y la conectividad no estandarizadas entre el hipotálamo, la amígdala y la cingulada anterior pueden generar sentimientos menos positivos (incluso negativos) acerca de las interacciones sociales. Lo que un sujeto de control podría interpretar como una interacción agradable —como cuando charla con un desconocido haciendo cola en una tienda o cuando ayuda a un vecino a cargar unas bolsas—, esos otros sujetos lo interpretan como interacciones desagradables. Son datos plausibles, debido a lo que ya sabemos sobre el importante papel que desempeña la amígdala en las sensaciones de miedo y de respuestas al miedo, así como en las respuestas sociales positivas.59 Son muchos los factores que ejercen una función en la sociabilidad de los seres humanos y —tal como debatiremos en el capítulo 5— los genes por sí solos rara vez surten grandes efectos, a no ser en función de si participan de redes genéticas de múltiples nódulos y de si participan de las redes de un entorno cerebral con múltiples bucles. Como consecuencia de ello, el hallazgo sobre una variante significativa del gen del OXTR, aunque es importante, seguramente sólo será parte de la historia de la sociabilidad humana y su amplia variabilidad.
Para llegar a apreciar estos otros factores, hay que tener en cuenta que existe un efecto generacional de la conducta materna en los niveles de OXT del bebé, así como en su consiguiente conducta social. Michael Meany y sus colaboradores demostraron que las ratas madre que registran niveles elevados de conducta materna también presentan niveles altos de OXT; los receptores de su conducta materna también tienen niveles altos de OXT y se ha demostrado que este hecho está en parte relacionado con el cuidado y las lameduras de la madre. Cuando los cachorros hembra maduran y tienen sus propias camadas, también ellas son muy maternales, también tienen niveles elevados de OXT y a su vez sus crías registran niveles elevados de OXT.60 Las pruebas cruzadas demuestran que la conducta de los progenitores en la experiencia temprana de las crías es más influyente en este asunto que los genes en sí mismos.61 Un resultado semejante se ha obtenido en el estudio de los macacos.62 En los seres humanos, los niveles elevados de OXT se corresponden con niveles altos de interacciones maternas, que a su vez se corresponden con niveles altos de OXT en los bebés. Tal y como Ruth Feldman y su equipo han indicado, se produce un bucle de biorretroalimentación entre la OXT, el cuidado de los hijos y la competencia social infantil en seres no humanos, aunque esta pauta también puede registrarse en humanos.63
¿HAY ALGO MÁS APARTE DE LA OXITOCINA?
Disponemos de más información acerca de la función que desempeña la vasopresina en machos que en hembras. En los machos resulta esencial para unirse a una pareja, y posiblemente también tenga algo que ver en las pautas agresivas, especialmente cuando son actos de defensa de las crías o de la pareja. No obstante, en condiciones concretas, la VPA surte efectos contrarios a los de la OXT. Por tanto, administrar VPA a un ratón macho incrementa su nivel de actividad y excitación, y eso suele relacionarse con posturas defensivas y no tanto con actitudes «amistosas». Mientras que administrar OXT a las hembras reduce su movilidad e induce quietud, la VPA administrada a los machos parece provocar el efecto contrario. Por supuesto, los sistemas de la OXT y la VPA interaccionan con otras hormonas como el estrógeno y la progesterona, tanto durante la gestación como después del nacimiento. También interaccionan con neurotransmisores como la dopamina y la serotonina,64 y actualmente se están estudiando los detalles. (Los neurotransmisores —que hay de muchos tipos— son sustancias secretadas por una neurona que se vinculan a otra neurona después de esparcirse en el espacio comprendido entre dos neuronas, constituyendo así una forma de comunicación entre neuronas que están separadas en el espacio. La sustancia liberada incrementará o reducirá la probabilidad de que la neurona receptora se active.)
Aparte de los sistemas de la OXT y la VPA, el sistema de la dopamina parece ser también importante en la expresión de la conducta social. La dopamina es un neurotransmisor que desempeña múltiples papeles en distintas funciones. Tiene dos tipos de receptor, el D1 y el D2,65 que son particularmente relevantes para la conducta social, y cada uno de ellos cumple una funcionalidad distinta. Se sabe que la dopamina es crucial para el aprendizaje, y que media en los cambios neuronales en el sistema de recompensa y castigo cuando los animales aprenden sobre el mundo y llegan a predecir un evento a partir de otro. Por ejemplo, para que los ratones de campo creen vínculos afectivos con su pareja, necesitan poder reconocer con qué otro ratón se han emparejado, ese reconocimiento requiere un aprendizaje, y ese aprendizaje requiere dopamina.
Desde hace poco sabemos que la dopamina desempeña una función en el afecto entre las parejas y en la conducta paternal/maternal. Se necesita tener acceso a los receptores de dopamina D2 para formar un vínculo afectivo de pareja, mientras que la activación de los receptores de dopamina D1 bloquean ese mismo vínculo. Después de la formación del vínculo, los receptores de D1 se regulan al alza, impidiendo así la formación de un segundo vínculo. Para que la dopamina funcione en los emparejamientos, sus receptores de D2 tienen que estar situados cerca de los receptores de la OXT en las mismas neuronas del sistema de recompensas; para las hembras, esa estructura de localización conjunta tiene que darse en el núcleo accumbens, mientras que en los machos, debe existir en el pálido ventral (ambas estructuras forman parte de los sistemas de recompensa y castigo).
La liberación de los opiáceos endógenos se produce después de la reunión de animales separados o de una respuesta satisfactoria a las llamadas de socorro de las crías.66 Por lo que a la conducta se refiere, puede observarse esta dinámica en la alegría que muestra un perro cuando se reúne con su compañero o con su amo, una conducta que es muy distinta de la tristeza que muestran cuando los amos dejan las maletas en el vestíbulo de su casa. Cuando los perros se reencuentran, por ejemplo, se lamerán la cara, darán saltos y moverán la cola con frenesí. La naturaleza exacta del papel que desempeñan los opiáceos endógenos, así como sus interacciones con otras hormonas como la prolactina, la OXT y la VPA, aún tiene que dilucidarse. Aunque las informaciones de las que disponemos son incompletas, y, además, se van complicando a medida que nos adentramos en este tema, su base fundamental (el hecho de que la densidad del receptor de OXT y VPA interviene en cada apego) explica parte del misterio.
Ya he señalado que existe una compleja relación entre la OXT y el FLC, una hormona del estrés, pero uno de los hallazgos más extraordinarios amplía esa complejidad hasta llevarla al ámbito inesperado de la salud en general y de la curación de heridas en particular. Las condiciones estresantes, como pasar apuros, pueden retrasar la cicatrización de heridas, un resultado que se da tanto en humanos como en roedores. Y lo que es aún más importante, se ha demostrado que si se administra OXT se acelera la cicatrización de heridas en ratas que padecen estrés. Este hallazgo suscita una pregunta muy inquietante acerca de las relaciones que existen entre la OXT y otras sustancias de las que se sabe que desempeñan un papel en la curación de heridas, como las citoquinas ambulantes (parte de la respuesta del sistema inmunológico) y otras sustancias que reducen la inflamación. En un artículo reciente, el neurocientífico Jean-Philippe Gouin y sus colaboradores67 hicieron pruebas sobre la cicatrización de heridas en sujetos humanos. Treinta y siete parejas, que mostraban muy distintos grados de afecto o de tensión, ingresaron para una visita hospitalaria de veinticuatro horas, y en ese período participaron en una «tarea de apoyo a la interacción social estructurada». Se midieron los niveles de OXT y de VPA en saliva cuando los sujetos ingresaron. Uno de los hallazgos fue que los niveles más altos de OXT y de VPA se relacionaban con las relaciones humanas afectuosas y de apoyo, y los niveles más bajos se relacionaban con una «comunicación negativa» entre los miembros de la pareja. En cuanto a la «herida», a todos los sujetos se les provocó una pequeña ampolla por succión en el antebrazo. El avance de la cicatrización de esa herida se supervisó a diario durante ocho días, y una vez más en el duodécimo día. Los principales hallazgos, dejando a un lado las estadísticas, fueron que los individuos con niveles altos de OXT cicatrizaban mucho más rápido, y las mujeres con niveles elevados de VPA también.
Asimismo, cabe señalar en este sentido las propuestas del uso terapéutico de la oxitocina en el tratamiento de los casos de trastorno por estrés postraumático (TEPT) que se resisten a la terapia cognitiva. Debido a la relación existente entre los niveles de oxitocina y las sensaciones de seguridad, confianza y placer que experimentamos en compañía de otras personas, y debido a que el debilitamiento de las respuestas condicionadas por el temor implica también un debilitamiento de las reacciones de la amígdala a un estímulo, la estrategia terapéutica de la oxitocina se está analizando.68
PATERNIDAD
Todo lo expuesto anteriormente nos permite situar parte de los datos conocidos sobre el afecto entre parejas, pero aún nos queda mucho por decir acerca de los mecanismos en virtud de los cuales los ratones macho de campo muestran una paternidad espontánea. Los nuevos datos indican que, en principio, la OXT y la VPA también intervienen en esta conducta, aunque no de modo exclusivo. La neurocientífica Karen Bales demostró que los ratones macho de campo sin experiencia reproductiva se implican en la alocría (cuidado de crías que no son propias) de camadas a las que están expuestos, mostrando así una paternidad pasiva (acariciar a las crías) y activa (apartarlos del peligro y lamerlos).69 Sin embargo, si se trata a los machos con sustancias que bloquean los receptores de OXT y de VPA, la alocría se reduce y los ataques contra los cachorros aumentan. Con dosis inferiores de bloqueantes, la respuesta a las crías es más lenta y los ataques, menores, lo cual indica una dependencia entre la reacción y la dosis. Si sólo se bloquea un tipo de receptor (tanto el OXTR como el VPAR), no se consigue el mismo efecto. Así pues, cualquiera de los dos receptores parece bastar para mediar en la alocría. Por último, parece ser que en un ratón macho de campo sin experiencia reproductiva, la simple exposición a las crías incrementa su nivel de OXT, reduce su nivel de corticostestora (una hormona del estrés) y refuerza las probabilidades de unirse posteriormente a una hembra.
Los biólogos evolucionistas se preguntarán por qué los ratones macho de campo se preocupan por cuidar de sus crías e incluso de crías que no son suyas. ¿Qué les aporta a ellos y a sus genes? A fin de cuentas, las camadas de ratón de montaña se las arreglan sin sus padres. Por lo que sé, no hay una respuesta definitiva a esta cuestión. Los entornos de los ratones de montaña y de campo son muy distintos, y los de campo son posiblemente más vulnerables a la depredación por parte de los halcones y los cernícalos que los ratones de montaña, que, por lo general, pueden resguardarse entre las rocas y los arbustos. En campo abierto, el cuidado de los machos puede ayudar a defender el nido, y, al aportar alimento adicional, los machos pueden fortalecer a sus crías para que sean más resistentes a la depredación. En cualquier caso, el hecho de que la mediación de la OXT y la VPA en la paternidad se aprecie en los machos también apunta hacia un concepto más global; a saber, que en el caso de los mamíferos se consigue una mayor sociabilidad con pocas modificaciones genéticas, lo cual a su vez modifica el circuito cerebral, las sustancias neuroquímicas y los receptores con el fin de favorecer nuevos niveles de sociabilidad.
Al inicio de las investigaciones en el emparejamiento monógamo de los ratones de campo, se decía que las diferencias genéticas entre las parejas monógamas y las demás podían relacionarse con las variantes de un tramo concreto del ADN que regula la expresión del receptor vasopresina. Se descubrió que ese tramo del ADN en concreto era más largo en los ratones de campo que en los de montaña, y por tanto se suscitó la pregunta de si eso se daría también en otras especies que se emparejan a largo plazo. Las investigaciones llevadas a cabo posteriormente en otras especies han cuestionado esta idea. En el emparejamiento a largo plazo intervienen múltiples mecanismos, y el análisis genético demuestra que las pautas de emparejamiento monógamo han evolucionado en múltiples ocasiones en los mamíferos, y al menos dos veces dentro del género del Peromyscus.70 No sería, por tanto, acertado referirnos a un «gen de la monogamia».71
¿Y qué hay de los vínculos afectivos que se establecen entre parejas humanas? ¿Somos nosotros, por naturaleza, como los ratones de campo? La respuesta a esta pregunta parece ser que los humanos son flexibles en sus pautas de apareamiento. Los vínculos intensos y a largo plazo son comunes, pero, según los antropólogos George Murdock y Suzanne Wilson, un 83% de las sociedades admiten patrones de poliginia. Sin embargo, según las circunstancias, aunque se permita la poliginia, la mayoría de hombres no disponen de suficientes recursos y por tanto optarán por tener una única esposa.72 En consecuencia, la monogamia de facto puede prevalecer, aunque los hombres ricos pueden tener más de una esposa. Los datos históricos nos indican que, en el pasado, un hombre rico podía establecer un vínculo afectivo especial a largo plazo con una hembra, aunque al mismo tiempo disfrutara de la compañía de otras mujeres e incluso las fecundara. Así pues, incluso en los casos en los que la poliginia es una práctica local, la tendencia de los individuos es la de formar vínculos afectivos a largo plazo.
En el 17% restante de sociedades, tanto modernas como antiguas (por ejemplo, Grecia y Roma), se ha practicado la monogamia. Una posible explicación a las variantes culturales que existen en las prácticas matrimoniales viene dada por las variaciones en las condiciones ecológicas y culturales, y, en concreto, por si existen convenciones para heredar propiedades y otras formas de riqueza, así como una fortuna específica que aguarda ser heredada.
Basándose en datos históricos y etnográficos, los biólogos Laura Fortunato y Marco Archetti arguyen que cuando un varón cuenta con varias esposas con hijos, y, por tanto, múltiples herederos, transferir los recursos a todos esos herederos da como resultado un debilitamiento de su valor como «aptos»; por ejemplo, las parcelas de tierra que se heredan son cada vez más pequeñas y pierden la capacidad de alimentar a las familias que dependen de esos terrenos.73 Un hombre podría seleccionar a una esposa en concreto cuyos hijos heredasen toda su fortuna, pero eso favorece la competitividad entre los miembros de la descendencia y suele ser una solución inestable. En estas condiciones, una estrategia más estable para reforzar el bienestar de la propia descendencia sería tener una única esposa, asegurarse de la paternidad de la descendencia e invertir concentradamente en el bienestar de los hijos de esa hembra. Fortunato y Archetti observan que la monogamia surgió en Eurasia a medida que la agricultura se iba extendiendo, ya que las tierras y los ganados se convirtieron en una fuente importante de riqueza que podía heredarse. Cuando ciertas prácticas se convierten en normativas, cuando se demuestra que aportan beneficios y reducen los problemas, y cuando se ven reforzadas por la aprobación y la desaprobación social, entonces todo indica que esas prácticas van en el camino correcto.
¿EN QUÉ CONSISTE LA RELACIÓN ENTRE EL APEGO AFECTIVO Y LA MORALIDAD?
Es probable que la OXT y la VPA, así como una amplia gama de distribuciones de receptores, sean elementos importantes para explicar los distintos estilos humanos de sociabilidad, y este dato neurobiológico acarrea ciertas implicaciones para el origen y la base de la moralidad humana. Los seres humanos, al igual que los babuinos, los marmosetas, los leones y algunos otros mamíferos, son seres intensamente sociables. Nuestros cerebros se estructuran para atender a nuestros propios intereses, pero también a los de nuestra prole. Aunque la vida social puede suponer muchas ventajas, también incrementa la competitividad entre el mismo grupo, así como la rivalidad para conseguir recursos entre parientes, parejas y vecinos. La resolución de problemas sociales, fundamentada en los vínculos afectivos, pero también moldeada por una preocupación por la reputación y el miedo a ser castigado o a ser excluido, provoca una cierta atenuación de los conflictos, como los que implican amenazan externas y rivalidades internas. Así pues, en los seres humanos, la monogamia como práctica social puede ser una buena solución para reducir la competitividad entre hembras, así como para heredar los recursos. Algunas soluciones sociales son más eficaces que otras, ya que permiten la estabilidad y la seguridad dentro del grupo, pero otras pueden ser socialmente inestables a largo plazo o pueden volverse inestables para el bienestar de los miembros cuando cambian las condiciones. La conducta social y la moral parecen ser parte del mismo espectro de acciones, en el sentido de que esas acciones que consideramos «morales» implican unos resultados de mayor trascendencia que las acciones de tipo social como hacer un regalo a una madre que acaba de dar a luz. El hecho de que la conducta social y moral forme parte de una misma línea continua está modestamente corroborado por los datos neurocientíficos que demuestran que tanto si un sujeto considera un acto como «social» como si lo considera «moral», las regiones de la corteza prefrontal que registran un incremento de la actividad son las mismas.74
En los seres humanos, las prácticas culturales, las convenciones y las instituciones van cambiando a medida que las soluciones a los problemas sociales se vuelven más complejas. Esas prácticas pueden ser muy explícitas, como aprender a no lamer un cuchillo en la mesa, o más implícitas, como aprender las formas aceptables de abrazar y besar a tu prole. Los seres humanos son extraordinariamente buenos aprendiendo, pero aún son mejores imitadores. A veces, sin darnos cuenta, copiamos gestos, estilos, tecnologías, prácticas y simbolismo grupal.
La resolución de problemas sociales es probablemente un ejemplo de resolución de problemas de ámbito general, y se fundamenta en la capacidad, que en algunos humanos resulta prodigiosa, de idear y evaluar las consecuencias de una acción planificada. También depende de la capacidad, posiblemente relacionada con la práctica del juego, de modificar las prácticas y las tecnologías actuales a medida que cambian las condiciones ambientales. La variabilidad cultural de las prácticas sociales de los humanos está bien documentada por los científicos de las ciencias sociales, y cubre una amplia gama que va desde la propiedad de las tierras o las normativas bancarias hasta las respuestas adecuadas a un insulto o a las formas más apropiadas de expresar el sentido del humor.75 Pero del mismo modo que existen coincidencias entre culturas en el adorno del cuerpo o en la cría de animales, también hay temas comunes sobre el castigo, la resolución de conflictos, las interacciones entre pareja e hijos, la gestión de la propiedad y la defensa grupal. Por ejemplo, el rechazo o evitación como forma de castigo a una conducta social inadecuada es un rasgo común en muchas culturas y especies, y la reconciliación después del conflicto suele implicar roces y caricias, así como una postura ritualizada de sumisión. El rechazo y la reconciliación se relacionan a su vez con cambios evidentes en el circuito neuronal que provocan dolor o alivio respectivamente.
La generalización de las prácticas sociales se debe en parte a la similitud de nuestros deseos sociales básicos y sus mecanismos neurobiológicos, que se conservan en todos los mamíferos con ciertas modificaciones según la especie.
Aún no es posible establecer con exactitud lo que el cerebro humano acepta o permite por acumulación cultural, y qué condiciones ecológicas lo refrendan.76
En el capítulo 4 prestaremos mayor atención a lo que podemos aprender de la neurociencia y la antropología sobre la extensión de la confianza y la cooperación más allá de los grupos familiares reducidos, hasta alcanzar a los parientes más lejanos e incluso a auténticos desconocidos.
En el caso de los mamíferos, podemos afirmar que existen numerosos procesos cerebrales que participan de las dinámicas de sociabilidad, pero son tres los factores que destacan en este sentido: (1) los impulsos para velar por nuestro bienestar y el de nuestra descendencia, pareja y afiliados; (2) la capacidad para evaluar y predecir lo que uno mismo y los demás sentirán y harán en determinadas circunstancias; y (3) un sistema neural de recompensas y castigos relacionados con la interiorización de prácticas sociales y su correcta aplicación, que está a su vez relacionado, en términos generales, con aprender las expectativas y las costumbres de los padres, los hijos y otros miembros de la familia.77
La forma que adopta la sociabilidad en los individuos de una misma especie depende de su posición y de cómo se ganan el sustento. La sociabilidad no se juega a «todo o nada», sino que viene escalonada. Los pumas tienden a ser muy poco sociables, los humanos tienden a serlo mucho, y los cuervos figuran en un estado intermedio. La sociabilidad depende también en gran medida de los recursos alimentarios. Tal y como Benjamin Kilham ha demostrado en sus maravillosos estudios, los osos negros, tildados desde siempre como criaturas solitarias a excepción de las manadas formadas por madres osas, serán asombrosamente sociales siempre y cuando existan reservas de comida para alimentar a todo el mundo.78 Por último, dentro de una misma especie se registra una amplia variación entre individuos, tal y como hemos observado anteriormente, y ello se debe en parte a los genes del receptor de la OXT, pero también a la interacción entre padres e hijos. Algunos seres humanos tienden a desarrollar una conducta grupal y se preocupan mucho por su reputación, mientras que otros viven a gusto en los márgenes de la sociedad y son felices con su excentricidad; en un extremo, encontramos a los seres humanos con trastornos claramente desfavorables para su sociabilidad, como el autismo.
Si los valores morales se afianzan en la neurobiología de la sociabilidad y si la cooperación es una conducta importante y moralmente relevante, el próximo paso de nuestra investigación es fijarnos con mayor atención en la cooperación, y por último discernir cómo es posible que las interacciones cooperativas en un clima de confianza puedan darse con asiduidad entre amigos y desconocidos sin ningún vínculo familiar. Al mismo tiempo, tenemos que ser conscientes de que la sociabilidad tiene también un lado oscuro, y, en el caso de los seres humanos, puede verdaderamente ser muy oscuro.