1
¿Qué es la vida?

Imperturbables noche y día, los radiotelescopios escudriñan los cielos. Cuarenta y dos de ellos se encuentran esparcidos en un grupo poco definido en la sierra cubierta de maleza del norte de California. Sus cuencos blancos parecen caras inexpresivas, todas dirigidas al unísono y esperanzadas hacia algún punto situado más allá del horizonte, como si se tratara de un punto de reunión para invasores extraterrestres que intentaran ir a casa. La incongruencia es apropiada. Los telescopios pertenecen al SETI,1 la búsqueda de inteligencia extraterrestre, una organización que ha estado explorando los cielos en busca de señales de vida desde hace medio siglo, infructuosamente. Incluso los protagonistas no son demasiado optimistas acerca de sus probabilidades de éxito; pero cuando la financiación se agotó hace algunos años, un llamamiento directo al público pronto hizo que el Conjunto de Telescopios Allen estuviera de nuevo operativo. Según mi opinión, la empresa es un símbolo conmovedor del inseguro sentido de la humanidad acerca de nuestro lugar en el universo, y en realidad de la fragilidad de la propia ciencia; tecnología de ciencia ficción tan inescrutable que parece omnisciencia, dedicada a un sueño tan ingenuo que apenas tiene base científica: que no estamos solos en el universo.
Incluso si el conjunto no consigue nunca detectar vida, sigue siendo valioso. Quizá no sea posible observar en el sentido opuesto a través de estos telescopios, pero este es su poder real. ¿Qué es exactamente lo que esperamos encontrar ahí afuera? ¿Acaso la vida en otros lugares del universo será tan parecida a nosotros que también emplea ondas de radio? ¿Pensamos que la vida en otros lugares tiene que estar basada en el carbono? ¿Necesitará agua? ¿Oxígeno? En realidad, estas preguntas no son acerca de la constitución de la vida en algún otro lugar del universo: se refieren a la vida en la Tierra, acerca de por qué la vida es de la manera como la conocemos. Estos telescopios son espejos, que reflejan estas preguntas a los biólogos terrestres. El problema es que la ciencia tiene que ver con predicciones. Las cuestiones más apremiantes de la física son acerca de por qué las leyes de la física son como son: ¿qué principios fundamentales predicen las propiedades conocidas del universo? La biología es menos predictiva, y no tiene leyes comparables a las de la física, pero aun así la capacidad predictiva de la biología evolutiva es embarazosamente mala. Sabemos muchísimas cosas sobre los mecanismos moleculares de la evolución y sobre la historia de la vida en nuestro planeta, pero muchas menos sobre qué partes de esa historia son azar (trayectorias que podrían haberse desarrollado de manera muy diferente en otros planetas) y qué pizcas están dictadas por las leyes o las limitaciones físicas.
Y ello no se debe a falta de esfuerzo. Este terreno es el patio en el que juegan premios Nobel jubilados y otras figuras señeras de la biología; pero a pesar de toda su sabiduría e intelecto, no consiguen empezar a ponerse de acuerdo entre ellos. Hace cuarenta años, en los albores de la biología molecular, el biólogo francés Jacques Monod escribió su famoso libro Le Hasard et la Nécéssité, que propugna de manera sombría que el origen de la vida en la Tierra fue un accidente anómalo, y que estamos solos en un universo vacío. Las líneas finales del libro se acercan a la poesía, una amalgama de ciencia y metafísica:
La antigua alianza está ya rota; el hombre sabe al fin que está solo en la inmensidad indiferente del universo, de donde surgió por azar. Igual que su destino, su deber no está escrito en ninguna parte. Puede escoger entre el reino arriba o la oscuridad abajo.
Desde entonces, otros han argumentado en el sentido opuesto: que la vida es un resultado inevitable de la química cósmica. Surgirá rápidamente, casi en todas partes. Una vez la vida medra en un planeta, ¿qué ocurre a continuación? De nuevo, no hay un consenso. Las limitaciones ingenieriles pueden obligar a la vida a seguir rutas convergentes hasta lugares parecidos, con independencia de dónde empezó. Debido a la gravedad, es probable que los animales que vuelen sean livianos y posean algo parecido a alas. En un sentido más general, puede ser necesario que la vida sea celular, compuesta de pequeñas unidades que mantengan sus entrañas diferentes del mundo exterior. Si tales restricciones son dominantes, la vida en otras partes puede parecerse mucho a la vida en la Tierra. Y al revés; quizá impere la contingencia: la estructura de la vida depende de los supervivientes aleatorios de accidentes globales como el impacto del asteroide que aniquiló a los dinosaurios. Hagamos retroceder el reloj hasta la época del Cámbrico, hace quinientos millones de años, cuando los animales aparecieron por primera vez y de forma explosiva en el registro fósil, y dejemos que de nuevo avance. ¿Sería este mundo paralelo semejante al nuestro? Quizá las colinas estarían llenas de gigantescos pulpos terrestres.
Una de las razones para disponer telescopios encarados hacia el espacio es que en la Tierra tratamos con un tamaño muestral de uno. Desde un punto de vista estadístico, no podemos decir qué limitó la evolución de la vida en la Tierra, si es que algo lo hizo. Pero si esto fuera cierto, no habría base para escribir este libro, o cualquier otro. Las leyes de la física se aplican en todo el universo, como hacen las propiedades y la abundancia de los elementos, de ahí la química plausible. La vida en la Tierra posee muchas propiedades extrañas que durante siglos han desafiado a la mente de los mejores biólogos: características tales como el sexo y el envejecimiento. Si a partir de primeros principios (a partir de la constitución química del universo) pudiéramos predecir por qué surgieron dichas características, por qué la vida es como es, entonces tendríamos otra vez acceso al mundo de la probabilidad estadística. La vida en la Tierra no es realmente una muestra de uno, sino que a efectos prácticos es una variedad infinita de organismos que evolucionan a lo largo de un tiempo infinito. Pero la teoría evolutiva no predice, a partir de primeros principios, por qué la vida en la Tierra tomó la dirección que tomó. Con ello no quiero decir que piense que la teoría evolutiva es errónea (no lo es), sino simplemente que no es predictiva. Mi argumento en este libro es que en realidad hay fuertes limitaciones a la evolución (limitaciones energéticas) que sí que hacen posible predecir algunos de los rasgos más fundamentales de la vida a partir de primeros principios. Antes de poder abordar estas limitaciones, hemos de considerar por qué la biología evolutiva no es predictiva, y por qué estas restricciones energéticas han pasado desapercibidas en gran medida; de hecho, por qué apenas nos hemos dado cuenta de que existe un problema. Únicamente en los últimos años ha resultado aparente sin ambages, y sólo para aquellos que siguen la biología evolutiva, que existe una discontinuidad profunda e inquietante en el meollo mismo de la biología.
Hasta cierto punto, podemos culpar al DNA de este lamentable estado de cosas. Irónicamente, puede decirse que la era moderna de la biología molecular, y de toda la extraordinaria tecnología del DNA que conlleva, se inició con un físico, específicamente con la publicación del libro de Erwin Schrödinger What is Life?2 en 1944. Schrödinger planteó dos puntos clave: primero, que de alguna manera la vida resiste la tendencia universal a la descomposición, el aumento de entropía (desorden) que estipula la segunda ley de la termodinámica; y segundo, que el truco de la evasión local que la vida hace de la entropía reside en los genes. Propuso que el material genético es un cristal «aperiódico», que no posee una estructura estrictamente repetitiva, y de ahí que pueda actuar como un «código», que según dicen es la primera vez que se utilizó el término en la literatura biológica. El propio Schrödinger suponía, junto con la mayoría de los biólogos de la época, que el cuasicristal en cuestión tenía que ser una proteína; pero al cabo de una década frenética, Crick y Watson habían inferido la estructura cristalina del propio DNA. En su segundo artículo en Nature en 1953, escribieron: «Por lo tanto, parece probable que la secuencia precisa de las bases sea el código que porta la información genética». Esta frase es la base de la biología moderna. En la actualidad, la biología es información, las secuencias de genomas se disponen in silico3 y la vida se define en términos de transferencia de información.
Los genomas son la puerta de entrada a un país encantado. Las resmas de código, 3.000 millones de letras en nuestro caso, se leen como una novela experimental, un relato ocasionalmente coherente en cortos capítulos entrecortados por bloques de texto repetitivo, versos, páginas en blanco, oleadas de conciencia: y una puntuación peculiar. Una proporción minúscula de nuestro propio genoma, menos del 2 %, codifica proteínas; una porción mayor es reguladora; y la función del resto es susceptible de causar broncas desmedidas entre científicos que de otro modo son educados.4 Esto aquí no tiene mayor importancia. Lo que es claro es que los genomas pueden codificar hasta decenas de miles de genes y una gran cantidad de complejidad regulatoria, capaz de especificar todo lo necesario para transformar una oruga en una mariposa o un niño en un humano adulto. La comparación de los genomas de animales, plantas, hongos y amebas unicelulares demuestra que entran en juego los mismos procesos. Podemos encontrar variantes de los mismos genes, los mismos elementos reguladores, los mismos replicadores egoístas (como los virus) y los mismos tramos de fruslerías repetitivas en genomas de tamaños y tipos enormemente diferentes. Las cebollas, el trigo y las amebas poseen más genes y más DNA que nosotros. Los anfibios, como las ranas y las salamandras, tienen tamaños de genoma que superan los dos órdenes de magnitud, y el genoma de algunas salamandras es 40 veces más largo que el nuestro, mientras que algunas ranas lo tienen de un tamaño que es menos de un tercio del nuestro. Si tuviéramos que resumir en una sola frase las limitaciones estructurales de los genomas, tendría que ser «cualquier cosa vale».
Esto es importante. Si los genomas son información, y no hay limitaciones importantes en el tamaño y la estructura del genoma, entonces tampoco hay limitaciones en la información. Esto no significa que no existan en absoluto limitaciones en los genomas. Es evidente que las hay. Las fuerzas que actúan sobre los genomas incluyen la selección natural, así como factores más aleatorios: duplicación accidental de genes, cromosomas o genomas enteros, inversiones, supresiones e invasiones de DNA parásito. El resultado de todo esto depende de factores tales como el nicho ecológico, la competencia entre especies y el tamaño de la población. Desde nuestro punto de vista, todos estos factores son impredecibles. Forman parte del ambiente. Si el ambiente está especificado de forma precisa, podremos llegar quizá a predecir el tamaño del genoma de una especie determinada. Pero un número infinito de especies viven en una variedad sin fin de microambientes, que van desde el interior de otras células hasta las ciudades humanas y hasta las profundidades presurizadas de los océanos. No es tanto «cualquier cosa vale» como «todo vale». Cabe esperar encontrar tanta variedad en los genomas como factores hay que actúan sobre ellos en estos ambientes diversos. Los genomas no predicen el futuro, sino que recuerdan el pasado: reflejan las exigencias de la historia.
Consideremos de nuevo otros mundos. Si la vida trata de información, y la información no está limitada, entonces no podemos predecir qué aspecto tendrá la vida en otro planeta, sólo que no contravendrá las leyes de la física. Tan pronto como ha surgido alguna forma de material hereditario, ya se trate de DNA o de otra cosa, entonces la trayectoria de la evolución se vuelve libre de restricciones por la información e impredecible a partir de primeros principios. Lo que aparezca realmente por evolución dependerá del ambiente exacto, de las contingencias de la historia y del ingenio de la selección. Pero volvamos a la Tierra. Esta afirmación es razonable para la enorme variedad de la vida tal como existe actualmente; pero, simplemente, no es verdad para la mayor parte de la larga historia de la Tierra. Durante miles de millones de años, parece que la vida estuvo limitada de maneras que no pueden interpretarse fácilmente en términos de genomas, historia o ambiente. Hasta hace poco, la peculiar historia de la vida en nuestro planeta estaba lejos de ser clara, e incluso en la actualidad existe mucho alboroto acerca de los detalles. Permítame el lector que esboce el panorama que está surgiendo, y que lo contraste con versiones más antiguas que ahora parecen ser erróneas.
Una breve historia de los primeros 2.000 millones de años de la vida
Nuestro planeta tiene unos 4.500 millones de años de antigüedad. Durante su historia inicial, durante unos 700 millones de años, estuvo arruinado por un duro bombardeo de asteroides, mientras el naciente sistema solar se asentaba. Un impacto colosal y temprano con un objeto del tamaño de Marte formó probablemente la Luna. A diferencia de la Tierra, cuya geología activa remueve continuamente la corteza, la superficie prístina de la Luna conserva pruebas de este bombardeo temprano en sus cráteres, que se han datado a partir de rocas que los astronautas del Apollo trajeron a la Tierra.
A pesar de la ausencia de rocas terrestres de edad comparable, existen todavía algunas pistas de las condiciones en la Tierra temprana. En particular, la composición de los circones (minúsculos cristales de silicato de circonio, de menor tamaño que granos de arena, que se encuentran en muchas rocas) sugiere que había océanos mucho antes de lo que se pensaba. Podemos decir a partir de la datación con uranio que algunos de estos cristales asombrosamente robustos se formaron hace entre 4.000 y 4.400 millones de años, y después se acumularon como granos detríticos en rocas sedimentarias. Los cristales de circón se comportan como minúsculas jaulas que atrapan contaminantes químicos, lo que refleja el ambiente en el que se formaron. La química de los primeros circones sugiere que se formaron a temperaturas relativamente bajas, y en presencia de agua. Lejos de la imagen de un infierno volcánico, con océanos de lava hirviente, que las impresiones de los artistas captaron vívidamente en lo que técnicamente se denomina el período «Hadeano», los cristales de circón indican un mundo acuático más tranquilo con una superficie terrestre limitada.
Asimismo, la antigua idea de una atmósfera primordial repleta de gases tales como metano, hidrógeno y amoníaco, no soporta el escrutinio de los circones. Elementos traza como el cerio se incorporan a los cristales de circón sobre todo en su forma oxidada. El elevado contenido de cerio en los circones más antiguos sugiere que la atmósfera estaba dominada por gases oxidados que emanaban de volcanes, en especial dióxido de carbono, vapor de agua, nitrógeno gas y dióxido de azufre. Esta mezcla no es diferente en composición a la del aire en la actualidad, excepto que faltaba el oxígeno, que no fue abundante hasta mucho más tarde, después de la aparición de la fotosíntesis. Leer la constitución de un mundo que hace mucho tiempo que desapareció a partir de unos cristales de circón, pocos y dispersos, pone mucho peso en lo que no dejan de ser granos de arena, pero esto es mejor que no disponer de prueba alguna. Dicha evidencia conjura de manera consistente un planeta que era sorprendentemente parecido al que conocemos hoy en día. Algún impacto ocasional de asteroides pudo haber vaporizado parcialmente los océanos, pero es improbable que hubiera afectado a cualesquiera bacterias que vivieran en los océanos profundos… si es que ya habían surgido por evolución.
Las primeras pruebas de vida son igualmente poco sólidas, pero pueden remontarse a algunas de las rocas más antiguas conocidas de Isua y Akilia, en el sudoeste de Groenlandia, que tienen unos 3.800 millones de años de antigüedad (véase la figura 2 para un cronograma). Estas pruebas no son en la forma de fósiles o de moléculas complejas derivadas de células vivas («biomarcadores»), sino que se trata simplemente de una ordenación no aleatoria de los átomos de carbono en el grafito. El carbono se encuentra en dos formas estables, o isótopos, que tienen masas marginalmente distintas.5 Los enzimas (proteínas que catalizan las reacciones en las células vivas) tienen una ligera preferencia por la forma más ligera, el carbono-12, que por lo tanto tiende a acumularse en la materia orgánica. El lector puede imaginar que los átomos de carbono son diminutas bolas de ping-pong: las bolas algo más pequeñas botan algo más rápidamente, de modo que es más probable que impacten en enzimas, y por ello es más probable que se conviertan en carbono orgánico. Por el contrario, la forma más pesada, el carbono-13, que constituye únicamente el 1,1 % del carbono total, tiene más probabilidades de quedarse en los océanos y puede en cambio acumularse cuando se precipita carbonato en rocas sedimentarias como la caliza. Estas minúsculas diferencias son consistentes hasta el extremo que a menudo se consideran diagnósticas de la vida. No sólo el carbono, sino otros elementos como el hierro, el azufre y el nitrógeno son fraccionados de manera parecida por las células vivas. Se ha informado de dicho fraccionamiento isotópico en las inclusiones de grafito en Isua y Akilia.
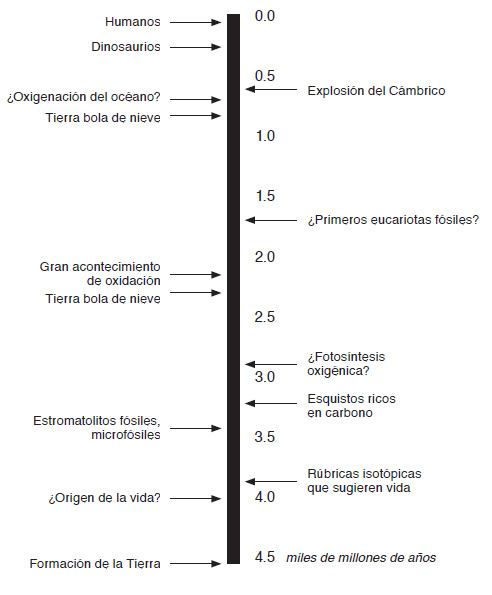
FIG. 2. Un cronograma de la vida. El cronograma muestra fechas aproximadas para algunos acontecimientos clave en la evolución temprana. Muchas de tales fechas son inciertas y abiertas al debate, pero la mayoría de las pruebas sugieren que bacterias y arqueos surgieron aproximadamente entre 1.500 y 2.000 millones de años antes que los eucariotas.
Se han puesto en tela de juicio todos los aspectos de este trabajo, desde la edad de las propias rocas hasta la existencia misma de los pequeños granos de carbono que se pretende que signifiquen presencia de seres vivos. Y todavía más: ha quedado claro que el fraccionamiento isotópico no es en absoluto algo único de la vida, sino que puede ser imitado, aunque de manera más débil, por procesos geológicos que tienen lugar en las fumarolas hidrotermales. Si las rocas de Groenlandia son realmente tan antiguas como parece, y si contienen ciertamente carbono fraccionado, esto no constituye todavía prueba de la existencia de vida. Esto puede parecer desalentador, pero en otro sentido no es menos de lo que cabría esperar. Argumentaré que la distinción entre un «planeta vivo» (un planeta geológicamente activo) y una célula viva es sólo una cuestión de definición. No existe una línea divisoria estricta. La geoquímica da paso a la bioquímica sin solución de continuidad. Desde este punto de vista, el hecho de que no podamos distinguir entre geología y biología en estas rocas antiguas es adecuado. Aquí tenemos un planeta vivo que da origen a la vida, y los dos no pueden separarse sin romper un continuo.
Si avanzamos unos pocos cientos de millones de años, la evidencia de la vida es más tangible: tan sólida y descifrable como las rocas antiguas de Australia y Sudáfrica. Allí existen microfósiles que tienen aspecto de células, aunque intentar situarlos en grupos modernos es una tarea ingrata. Muchos de estos fósiles minúsculos están revestidos de carbono, que de nuevo presenta rúbricas isotópicas delatoras, pero que ahora son algo más consistentes y pronunciadas, lo que sugiere un metabolismo organizado en lugar de procesos hidrotermales casuales. Y hay estructuras que parecen estromatolitos, estas catedrales abovedadas de vida bacteriana, en las que las células crecen capa sobre capa, las capas enterradas se mineralizan y se transforman en piedra, y en último término construyen estructuras rocosas asombrosamente laminadas, de un metro de alto. Más allá de estos fósiles directos, hace 3.200 millones de años hay características geológicas a gran escala, de cientos de kilómetros cuadrados de superficie y de decenas de metros de potencia, notablemente las formaciones de hierro bandeado y los esquistos ricos en carbono. Solemos pensar que las bacterias y los minerales ocupan reinos diferentes, lo vivo frente a lo inanimado, pero de hecho muchas rocas sedimentarias se depositan, a una escala colosal, mediante procesos bacterianos. En el caso de las formaciones de hierro bandeado (que son de una belleza pasmosa, en sus bandas de rojo y negro), las bacterias extraen electrones del hierro disuelto en el océano (este hierro «ferroso» es abundantísimo en ausencia de oxígeno) y dejan atrás el pellejo insoluble, la herrumbre, que se hunde hasta las profundidades. Sigue siendo un misterio por qué estas rocas ricas en hierro son listadas, pero aquí también la rúbrica isotópica delata la mano de la biología.
Estos enormes depósitos indican no sólo vida, sino fotosíntesis. No la forma familiar de fotosíntesis que vemos a nuestro alrededor en las hojas verdes de las plantas y en las algas, sino un precursor más sencillo. En todas las formas de la fotosíntesis, la energía de la luz se emplea para arrancar electrones de un donante renuente. Después los electrones son introducidos en el dióxido de carbono para formar moléculas orgánicas. Las diversas formas de fotosíntesis difieren en su fuente de electrones, que pueden proceder de todo tipo de lugares distintos, siendo los más comunes el hierro disuelto (ferroso), el sulfuro de hidrógeno o el agua. En cada caso, los electrones son transferidos al dióxido de carbono, y dejan atrás los desechos: depósitos de hierro oxidado, azufre elemental (alcrebite) y oxígeno, respectivamente. La nuez más difícil de cascar, con mucho, es el agua. Hace 3.200 millones de años la vida extraía electrones de casi todo lo demás. La vida, como observó el biólogo Albert Szent-Györgyi, no es otra cosa que un electrón en busca de un lugar para descansar. Es motivo de debate el momento exacto en el que tuvo lugar el paso final de extraer electrones del agua. Hay quien dice que fue un acontecimiento temprano en la evolución, pero el peso de las pruebas sugiere ahora que la fotosíntesis «oxigénica» surgió hace entre 2.900 y 2.400 millones de años, no mucho antes de un período cataclísmico de agitación global, la crisis de mediana edad de la Tierra. Extensas glaciaciones globales, conocidas como la «Tierra bola de nieve», fueron seguidas por la oxidación generalizada de las rocas terrestres, hace unos 2.200 millones de años, que dejaron «lechos rojos» oxidados como señal definitiva de presencia de oxígeno en el aire: el «Gran Evento de Oxidación». Incluso las glaciaciones globales indican un aumento del oxígeno atmosférico. Al oxidar el metano, el oxígeno retiró del aire un potente gas de invernadero, desencadenando la congelación global.6
Con la evolución de la fotosíntesis oxigénica, la caja de herramientas metabólicas de la vida estaba esencialmente completa. Es improbable que nuestro recorrido con paradas programadas de cerca de dos mil millones de años de historia de la Tierra (tres veces más extenso que toda la duración de los animales) sea exacto en todos sus detalles, pero será bueno hacer una breve pausa para considerar qué es lo que el panorama general dice acerca de nuestro mundo. Primero, la vida surgió muy pronto, probablemente hace entre 3.500 y 4.000 millones de años, si no antes, en un mundo acuático no distinto del nuestro. Segundo, hace entre 3.500 y 3.200 millones de años, las bacterias ya habían inventado la mayoría de las formas del metabolismo, incluidas múltiples formas de respiración y de fotosíntesis. Durante mil millones de años, el mundo fue un caldero de bacterias, que exhibían una inventiva de bioquímica que apenas podemos imaginar.7 El fraccionamiento isotópico sugiere que todos los ciclos principales de nutrientes (carbono, nitrógeno, azufre, hierro, etc.) ya estaban en marcha antes de hace 2.500 millones de años. Pero sólo con el aumento del oxígeno, desde hace 2.400 millones de años, transfiguró la vida nuestro planeta hasta el punto de que este mundo bacteriano próspero podría haber sido detectado como un planeta vivo desde el espacio. Sólo entonces empezó la atmósfera a acumular una mezcla reactiva de gases, como el oxígeno y el metano, que las células vivas reponen continuamente, lo que delata la mano de la biología a una escala planetaria.
El problema con los genes y el ambiente
Hace tiempo que se reconoce que el Gran Evento de Oxidación fue un momento crucial en la historia de nuestro planeta vivo, pero en los últimos años su importancia ha cambiado de forma radical, y la nueva interpretación es fundamental para mi argumentación en este libro. La versión antigua considera que el oxígeno fue el determinante ambiental crítico para la vida. El oxígeno no especifica qué es lo que evolucionará, indica la argumentación, pero permite la evolución de una complejidad mucho mayor: suelta el freno. Los animales, por ejemplo, se ganan la vida moviéndose físicamente, persiguiendo presas o siendo perseguidos. Obviamente, ello requiere una gran cantidad de energía, de modo que es fácil imaginar que los animales no podrían existir en ausencia de oxígeno, que proporciona casi un orden de magnitud más de energía que otras formas de respiración.8 Esta afirmación es tan sosa y poco interesante que ni siquiera vale la pena ponerla en entredicho. Esto es parte del problema: no invita a una consideración ulterior. Podemos dar por sentado que los animales necesitan oxígeno (aunque esto no es siempre cierto), y lo tienen, de modo que el oxígeno es un común denominador. Los problemas reales en biología evolutiva tienen que ver pues con las propiedades y el comportamiento de los animales o las plantas. O esto es lo que parece.
Esta concepción respalda implícitamente la historia de la Tierra que aparece en los manuales. Solemos pensar que el oxígeno es saludable y bueno, pero en realidad, desde el punto de vista de la bioquímica primordial, no lo es en absoluto: es tóxico y reactivo. A medida que los niveles de oxígeno aumentaron, cuenta el relato de los libros de texto, este gas peligroso supuso una fuerte presión de selección para todo el mundo microbiano. Hay relatos crueles de la extinción en masa que acabó con todos ellos, lo que Lynn Margulis denominó el «holocausto del oxígeno». El hecho de que no haya trazas de dicho cataclismo en el registro fósil no debería preocuparnos mucho (se nos asegura): estos bichos eran muy pequeños y todo aquello ocurrió hace mucho, muchísimo tiempo. El oxígeno obligó a nuevas relaciones entre las células: simbiosis y endosimbiosis, en las que las células intercambiaron entre ellas y dentro de ellas útiles de supervivencia. A lo largo de cientos de millones de años, la complejidad aumentó gradualmente, a medida que las células aprendían no sólo a habérselas con el oxígeno, sino también a aprovecharse de su reactividad: desarrollaron la respiración aerobia, que les confería mucha más energía. Estas células grandes, complejas y aerobias empaquetan su DNA en un compartimento especial, llamado núcleo, lo que se refleja en su nombre, «eucariotas»: literalmente, «de núcleo verdadero». Reitero que este es un relato de manual; argumentaré que es erróneo.
Hoy en día, todos los seres vivos complejos que vemos a nuestro alrededor, todas las plantas, animales, algas, hongos y protistas (células grandes, como las amebas), están compuestos de estas células eucariotas. Los eucariotas aumentaron progresivamente hasta dominar a lo largo de mil millones de años, continúa el relato, en un período conocido, irónicamente, como los «aburridos mil millones», pues en el registro fósil ocurrieron pocas cosas. Aun así, entre hace 1.600 y 1.200 millones de años empezamos a encontrar fósiles de unicelulares que se parecen mucho a eucariotas, y algunos de los cuales incluso encajan cómodamente en grupos modernos, como las algas rojas y los hongos.
Después vino otro período de agitación global y una sucesión de Tierras bola de nieve hace unos 750-600 millones de años. Poco después, los niveles de oxígeno aumentaron rápidamente hasta cerca de los niveles modernos; y los primeros fósiles de animales aparecen abruptamente en el registro fósil. Los primeros fósiles grandes, de hasta un metro de diámetro, son un grupo misterioso de formas con aspecto de frondes que la mayoría de los paleontólogos interpretan como animales filtradores, aunque algunos insisten en que son simplemente líquenes: los fósiles ediacarianos9 o, de manera más cariñosa, vendobiontes. Después, tan abruptamente como aparecieron, la mayoría de estas formas desaparecieron en una extinción en masa propia, para ser sustituidas, en los albores de otra era, el Cámbrico, hace 541 millones de años (una fecha tan emblemática entre los biólogos como 106610 o 1492), por una explosión de animales más reconocibles. Grandes y móviles, con ojos complejos y apéndices inquietantes, estos feroces depredadores y sus temibles presas acorazadas irrumpieron en la escena evolutiva, con colmillos y garras rojos, Darwin con aspecto moderno.11
¿Qué parte de este escenario hipotético es en realidad errónea? A primera vista parece ser plausible. Pero en mi opinión el subtexto está equivocado; y cuántas más cosas sabemos, también lo están muchos de los detalles. El subtexto está relacionado con la interacción entre los genes y el ambiente. Todo el escenario gira alrededor del oxígeno, que supuestamente es la variable ambiental clave, que permitió el cambio genético, al dejar de presionar el freno a la innovación. Los niveles de oxígeno aumentaron dos veces, en el Gran Evento de Oxidación hace 2.400 millones de años, y otra vez hacia el final del eterno período Precámbrico, hace 600 millones de años (figura 2). Cada vez, según el relato clásico, el aumento de oxígeno liberó de limitaciones a estructura y función. Después del Gran Evento de Oxidación, con sus nuevas amenazas y oportunidades, las células intercambiaron entre ellas en una serie de endosimbiosis, y acumularon gradualmente la complejidad de las verdaderas células eucariotas. Cuando los niveles de oxígeno aumentaron por segunda vez, antes de la explosión del Cámbrico, se dejaron de lado por completo las limitaciones físicas, como por arte de magia, y revelaron por primera vez la posibilidad de animales. Nadie afirma que el oxígeno impulsó físicamente estos cambios; más bien, transformó el paisaje selectivo. A lo largo de las magníficas vistas panorámicas de este nuevo paisaje sin limitaciones, los genomas se expandieron libremente, al hallarse finalmente sin trabas su contenido en información. La vida floreció y llenó todos los nichos concebibles, de todas las maneras posibles.
Esta concepción de la evolución puede considerarse en términos del materialismo dialéctico, fiel a los principios de algunos biólogos evolutivos importantes durante la síntesis neodarwiniana de principios a mediados del siglo XX. Los opuestos que se interpenetran son genes y ambiente, también conocidos como naturaleza y alimentación. La biología va toda de genes, y su comportamiento va todo del ambiente. Después de todo, ¿qué otra cosa hay? Bueno, la biología no va sólo de genes y ambiente, sino también de células y de las limitaciones de su estructura física, que veremos que tiene poco que ver directamente ni con los genes ni con el ambiente. Las predicciones que surgen de estas visiones dispares del mundo son asombrosamente distintas.
Tomemos la primera posibilidad, interpretar la evolución en términos de los genes y el ambiente. La falta de oxígeno en la Tierra primordial es una limitación ambiental fundamental. Añadamos oxígeno y la evolución florece. Toda la vida que está expuesta al oxígeno es afectada de una manera u otra y ha de adaptarse. Algunas células resultan ser más adecuadas para las condiciones aeróbicas, y proliferan; otras mueren. Pero hay muchos microambientes distintos. El aumento de oxígeno no inunda simplemente todo el mundo con oxígeno en una especie de ecosistema global monomaníaco, sino que oxida minerales en tierra y solutos en los océanos, y que también enriquece nichos anaerobios. Aumenta la disponibilidad de nitrato, nitrito, sulfato, sulfito, etc. Todos ellos pueden ser usados en lugar del oxígeno en la respiración celular, de modo que la respiración anaerobia medra en un mundo aerobio. Todo esto se resume en muchas maneras diferentes de ganarse la vida en el nuevo mundo.
Imagine el lector una mezcla aleatoria de células en un ambiente. Algunas células, como las amebas, viven a base de englobar físicamente a otras células, un proceso denominado fagocitosis. Algunas son fotosintéticas. Otras, como los hongos, digieren su alimento externamente: osmotrofia. Suponiendo que la estructura celular no imponga restricciones insuperables, haríamos la predicción de que estos tipos diferentes de células descienden de varios antepasados bacterianos diferentes. Resultó que una célula ancestral era algo mejor en alguna forma primitiva de fagocitosis, otra en una forma sencilla de osmotrofia, otra en la fotosíntesis. A lo largo del tiempo, sus descendientes se hicieron más especializados y mejor adaptados a este modo de vida concreto.
Planteémoslo de manera más formal: si los niveles crecientes de oxígeno permitieron que medraran nuevos estilos de vida, esperaríamos ver una radiación polifilética, en la que células u organismos no relacionados (de tipos diferentes) se adaptaran de forma célere, radiando nuevas especies que llenaran nichos vacíos. Este tipo de pauta es exactamente la que vemos… a veces. En la explosión del Cámbrico, por ejemplo, radiaron docenas de diferentes tipos animales, desde las esponjas a los equinodermos, desde los artrópodos a los gusanos. Estas grandes radiaciones animales estuvieron acompañadas de radiaciones equivalentes entre las algas y los hongos, así como entre los protistas, como los ciliados. La ecología se hizo enormemente compleja, y esto por sí mismo impulsó otros cambios. Fuera o no específicamente la marea creciente de oxígeno lo que desencadenó la explosión del Cámbrico, existe un consenso general de que los cambios ambientales transformaron efectivamente la selección. Algo ocurrió, y el mundo cambió para siempre.
Contrastemos esta pauta con lo que cabría ver si hubieran dominado las restricciones de estructura. Hasta que la restricción se supera, veríamos un cambio limitado en respuesta a cualesquiera variaciones ambientales. Cabría esperar largos períodos de estasis, insensibles a los cambios ambientales, con radiaciones monofiléticas muy ocasionales. Esto quiere decir que si, en una rara ocasión, un grupo concreto supera sus limitaciones estructurales intrínsecas, sólo este radiará para llenar nichos vacíos (aunque posiblemente con demora, hasta que un cambio en el ambiente lo permita). Desde luego, también vemos esto. En la explosión del Cámbrico vemos la radiación de diferentes grupos animales… pero no múltiples orígenes de animales. Todos los grupos animales comparten un antepasado común, como de hecho hacen también todas las plantas. El desarrollo multicelular12 complejo, que implica una línea germinal y un soma (cuerpo) distintos, es difícil. Aquí las limitaciones están relacionadas en parte con los requisitos de un programa de desarrollo preciso, que ejerce un férreo control sobre el destino de las células individuales. Pero a un nivel más laxo, un cierto grado de desarrollo multicelular es común, con hasta treinta orígenes separados de pluricelularidad entre grupos que incluyen las algas, los hongos y los mohos mucilaginosos. Pero hay un lugar en el que parece que las limitaciones de la estructura física (estructura celular) dominan hasta un grado tal que superan todo lo demás: el origen de la célula eucariota (células grandes y complejas) a partir de bacterias, como secuela del Gran Evento de Oxidación.
El agujero negro en el meollo de la biología
Si las células eucariotas complejas evolucionaron realmente en respuesta al aumento de oxígeno atmosférico, cabría predecir una radiación polifilética, con varios grupos diferentes de bacterias que engendraran de manera independiente tipos celulares más complejos. Cabría esperar ver que las bacterias fotosintéticas dieran origen a algas mayores y más complejas, que las bacterias osmotróficas dieran origen a los hongos, y las células móviles depredadoras a los fagocitos, y así sucesivamente. Esta evolución de mayor complejidad podría ocurrir mediante mutaciones genéticas estándar, intercambio génico y selección natural, o mediante incorporaciones y adquisiciones de endosimbiosis, tal como concibió Lynn Margulis en la bien conocida teoría de la endosimbiosis serial. De un modo u otro, si no hay limitaciones fundamentales en la estructura celular, entonces los niveles crecientes de oxígeno tendrían que haber hecho posible una mayor complejidad con independencia de cómo esta hubiera surgido exactamente por evolución. Predeciríamos que el oxígeno liberaría las limitaciones en todas las células, permitiendo una radiación polifilética con todo tipo de bacterias diferentes haciéndose más complejas de manera independiente. Pero no es esto lo que vemos.
Permítame el lector que lo explique con más detalle, pues el razonamiento es fundamental. Si las células complejas surgieron mediante selección natural «estándar», en la que las mutaciones genéticas dan origen a variaciones sobre las que actúa la selección natural, entonces cabría esperar ver todo un surtido de estructuras internas, tan variadas como el aspecto externo de las células. Las células eucariotas son maravillosamente variadas en su forma y tamaño, desde células algales gigantes, de aspecto de hoja, hasta neuronas alargadas y amebas extendidas. Si los eucariotas hubieran desarrollado la mayor parte de su complejidad en el curso de la adaptación a distintos modos de vida en poblaciones divergentes, entonces esta larga historia tendría que reflejarse también en sus estructuras internas distintivas. Pero si miramos en su interior (como haremos pronto), veremos que todos los eucariotas están hechos básicamente de los mismos componentes. La mayoría de nosotros no podríamos distinguir entre una célula vegetal, una célula renal y un protista procedente de la charca local si los situamos todos bajo el microscopio electrónico: todos tienen un aspecto notablemente parecido. Pruébelo el lector (figura 3). Si el aumento de los niveles de oxígeno eliminó las restricciones a la complejidad, la predicción a partir de la selección natural «estándar» es que la adaptación a diferentes modos de vida en diferentes poblaciones habría de conducir a la radiación polifilética. Pero no es esto lo que vemos.
Desde finales de la década de 1960, Lynn Margulis aducía que esta teoría era, en cualquier caso, equivocada: que las células eucariotas no surgieron mediante selección natural estándar, sino a través de una serie de endosimbiosis, en las que varias bacterias cooperaron juntas tan estrechamente que algunas células penetraron físicamente dentro de otras. Tales ideas se remontan a principios del siglo XX, a Richard Altmann, Konstantin Mereschkowski, George Portier, Ivan Wallin y otros, que sostenían que todas las células complejas surgieron mediante simbiosis entre células más sencillas. Sus ideas no fueron olvidadas, pero sí ridiculizadas y desechadas como «demasiado fantásticas para mencionarlas ahora en la sociedad biológica educada». En la época de la revolución de la biología molecular de la década de 1960, Margulis disponía de una base más firme, aunque todavía polémica, y ahora sabemos que al menos dos de los componentes de las células eucariotas proceden de bacterias endosimbiontes: las mitocondrias (los transductores de energía en las células complejas), que derivan de α-proteobacterias; y los cloroplastos (la maquinaria fotosintética de las plantas), que proceden de cianobacterias. Asimismo, en uno u otro momento, casi todos los demás «orgánulos» especializados de las células eucariotas han sido considerados endosimbiontes, entre ellos el propio núcleo, los cilios y flagelos (extensiones sinuosas cuyo batido rítmico impulsa el movimiento de las células) y los peroxisomas (fábricas del metabolismo tóxico). Así, la teoría de la endosimbiosis serial afirma que los eucariotas están compuestos de un conjunto de bacterias, forjadas en una empresa común a lo largo de cientos de millones de años después del Gran Evento de Oxidación.
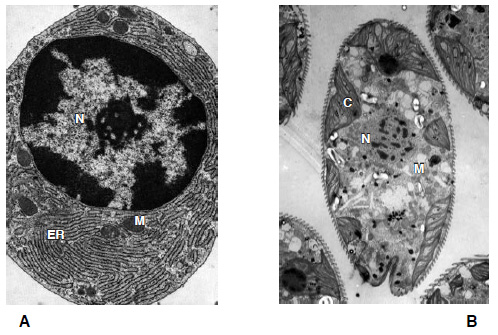
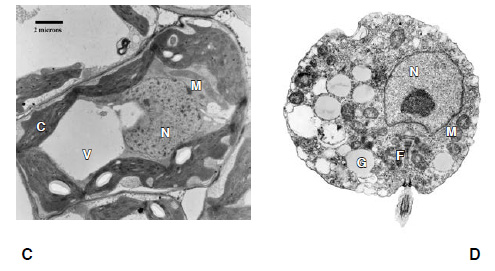
FIG. 3. La complejidad de los eucariotas. Cuatro células eucariotas diferentes que muestran una complejidad morfológica equivalente. A muestra una célula animal (una célula del plasma), con un gran núcleo central (N), membranas internas extensas (retículo endoplasmático, ER) tachonadas de ribosomas, y mitocondrias (M). B es el alga unicelular Euglena, que se encuentra en muchas charcas, que muestra un núcleo central (N), cloroplastos (C) y mitocondrias (M). C es una célula vegetal limitada por una pared celular, con una vacuola (V), cloroplastos (C), un núcleo (N) y mitocondrias (M). D es una zoospora de un hogo quítrido, implicado en la extinción de 150 especies de ranas; se aprecia el núcleo (N), mitocondrias (M), el flagelo (F) y cuerpos gamma (G) de función desconocida.
Se trata de una idea poética, pero la teoría de la endosimbiosis serial efectúa una predicción implícita equivalente a la de la selección estándar. Si fuera cierta, cabría esperar ver orígenes polifiléticos: un surtido de estructuras internas, tan variadas como el aspecto externo de las células. En cualquier serie de endosimbiosis, en la que la simbiosis dependa de algún tipo de intercambio metabólico en un ambiente concreto, cabría esperar encontrar tipos dispares de células que interactuaran en ambientes diferentes. Si tales células posteriormente se modelaran en los orgánulos de las células eucariotas complejas, la hipótesis predice que algunos eucariotas tendrían que poseer un conjunto de componentes, y otros un conjunto distinto. Cabría esperar encontrar todo tipo de situaciones intermedias y de variantes no relacionadas acechando en lugares escondidos y oscuros, como fangos pútridos. Hasta su muerte prematura debida a un derrame cerebral en 2011, Margulis se mantuvo efectivamente firme en su creencia de que los eucariotas son un tapiz rico y variado de endosimbiosis. Para ella, la endosimbiosis era un modo de vida, una vía «femenina» inexplorada de la evolución, en la que la cooperación («trabajar en red», como ello la llamaba) superaba la competencia desagradablemente masculina entre los cazadores y los cazados. Pero en su veneración de las células vivas «reales», Margulis volvió la espalda a la disciplina informática más árida de la filogenia, el estudio de las secuencias génicas y de los genomas completos, que tiene el poder de decirnos exactamente cómo eucariotas diferentes se relacionan entre sí. Y que cuenta un relato muy diferente, y en último término mucho más convincente.
El relato depende de un gran grupo de especies (un millar o más en número) de eucariotas unicelulares que carecen de mitocondrias. Antaño se consideraba que este grupo era un primitivo «eslabón perdido» evolutivo entre las bacterias y los eucariotas más complejos, exactamente el tipo de elemento intermedio que predecía la teoría de la endosimbiosis serial. El grupo incluye el molesto parásito intestinal Giardia, que en palabras de Ed Yong se parece a una lágrima malévola (figura 4). Giardia vive haciendo honor a su aspecto, causando desagradables diarreas. No tiene un único núcleo, sino dos, de modo que es incuestionable que es un eucariota, pero carece de otros rasgos arquetípicos, notablemente de mitocondrias. A mediados de la década de 1980, el iconoclasta biólogo Tom Cavalier-Smith proponía que Giardia y otros eucariotas relativamente simples eran probablemente los supervivientes del período más antiguo de la evolución de los eucariotas, antes de la adquisición de las mitocondrias. Aunque Cavalier-Smith aceptaba que las mitocondrias derivan efectivamente de endosimbiontes bacterianos, tenía poco tiempo para dedicar a la teoría de la endosimbiosis serial de Margulis; en lugar de ello, presentaba (y todavía lo hace) a los primeros eucariotas como fagocitos primitivos, similares a las modernas amebas, que obtenían su sustento englobando otras células. Las células que adquirieron mitocondrias, argumentaba, ya poseían un núcleo, y un esqueleto interno dinámico que les ayudaba a cambiar de forma y a desplazarse, y maquinaria proteínica para mover materiales por su interior, y compartimentos especializados para digerir alimento internamente, etc. Adquirir mitocondrias facilitaba las cosas, ciertamente: equiparon con turbo a estas células primitivas. Pero trucar un automóvil no altera la estructura del mismo: se sigue empezando con un automóvil, que ya posee un motor, cambio de marchas, frenos, todo lo que hace que sea un coche. Equipar con turbo no cambia nada excepto la potencia de salida. Lo mismo ocurre en el caso de los fagocitos primitivos de Cavalier-Smith: todo estaba ya en su lugar excepto las mitocondrias, que simplemente proporcionaron más energía a las células. Si hay una imagen de manual de los orígenes de los eucariotas (incluso en la actualidad) es esta.
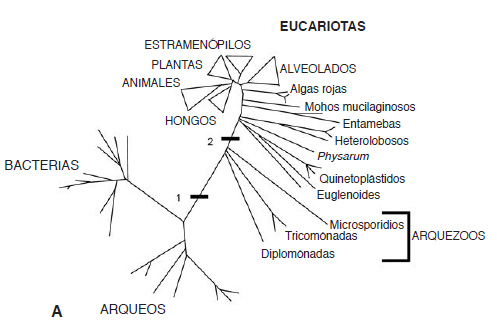
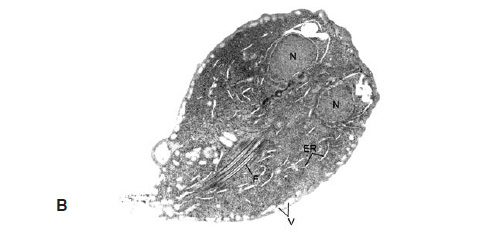
FIG. 4. Los arquezoos, el mítico (pero falso) eslabón perdido. A Un árbol de la vida antiguo pero engañoso basado en el RNA ribosómico, que muestra los tres reinos de bacterias, arqueos y eucariotas. Las barras marcan la supuesta evolución temprana del núcleo (1) y la supuesta adquisición tardía de las mitocondrias (2). Los tres grupos que se ramifican entre las barras constituyen los arquezoos, eucariotas presuntamente primitivos que todavía no habían adquirido mitocondrias, como Giardia (B). Ahora sabemos que los arquezoos no son en absoluto eucariotas primitivos, sino que proceden de antepasados más complejos que ya poseían mitocondrias; en realidad, surgen de la parte principal del árbol eucariota (N = núcleo; ER = retículo endoplasmático; V = vacuolas; F = flagelos).
Cavalier-Smith llamó a estos eucariotas primitivos «arquezoos» (que significa animales antiguos) para reflejar su supuesta antigüedad (figura 4). Varios de ellos son parásitos que causan enfermedades, de modo que su bioquímica y sus genomas han atraído el interés de la investigación médica y la financiación que ella conlleva. A su vez, esto significa que ahora sabemos muchas cosas acerca de ellos. A lo largo de las dos últimas décadas, hemos descubierto a través de sus secuencias genómicas y de su bioquímica detallada que ninguno de los arquezoos es un eslabón perdido real, lo que quiere decir que no son verdaderas formas evolutivas intermedias. Por el contrario, todos ellos derivan de eucariotas más complejos, que antaño poseían una cuota completa de todo, incluidas, y en particular, mitocondrias. Perdieron su antigua complejidad mientras se especializaban para vivir en nichos más simples. Todos ellos conservan estructuras que ahora se sabe que derivan de mitocondrias por una evolución reductiva, ya sea hidrogenosomas o mitosomas. Estas no se parecen mucho a mitocondrias, aunque tienen una estructura equivalente de membrana doble, de ahí la suposición errónea de que los arquezoos no tuvieron nunca mitocondrias. Pero la combinación de datos moleculares y filogenéticos demuestra que hidrogenosomas y mitosomas proceden en realidad de mitocondrias, no de algunos otros endosimbiontes bacterianos (como había predicho Margulis). Así, todos los eucariotas poseen mitocondrias de una forma u otra. Podemos inferir que el último antepasado común eucariota ya tenía mitocondrias, tal como había predicho Bill Martin en 1998 (véase la Introducción). El hecho de que todos los eucariotas posean mitocondrias puede parecer un punto trivial, pero cuando se combina con la proliferación de secuencias de genomas procedentes del mundo microbiano, más amplio, este conocimiento ha hecho cambiar completamente nuestra comprensión de la evolución de los eucariotas.
Ahora sabemos que todos los eucariotas comparten un antepasado común, que por definición surgió sólo una vez en los 4.000 millones de años de vida sobre la Tierra. Permítaseme reiterar este punto, pues es crucial. Todas las plantas, animales, algas, hongos y protistas comparten un antepasado común: los eucariotas son monofiléticos. Esto significa que las plantas no evolucionaron a partir de un tipo de bacterias, y los animales o los hongos a partir de otros tipos. Por el contrario, una población de células eucariotas morfológicamente complejas surgió en una única ocasión, y todas las plantas, animales, algas y hongos evolucionaron a partir de esta población fundadora. Cualquier antepasado común es, por definición, una entidad singular; no una célula única, sino una población única de células esencialmente idénticas. Esto no significa en sí mismo que el origen de las células complejas fuera un acontecimiento raro. En principio, las células complejas pudieron haber surgido en numerosas ocasiones, pero sólo persistió un grupo: todos los demás se extinguieron por alguna razón. Razonaré que no es este el caso, pero primero hemos de considerar las propiedades de los eucariotas con algo más de detalle.
El antepasado común de todos los eucariotas dio pronto origen a cinco «supergrupos» con diversas morfologías celulares, la mayoría de las cuales resultan desconocidas incluso para los biólogos de formación clásica. Dichos supergrupos tienen nombres tales como unicontos (que comprenden animales y hongos), excavados, cromalveolados y plantas (que incluyen las plantas terrestres y las algas). Sus nombres no importan, pero hay dos aspectos importantes. Primero, existe mucha más variación genética en el seno de cada uno de estos supergrupos que la que hay entre los antepasados de cada grupo (figura 5). Esto implica una radiación temprana y explosiva, específicamente una radiación monofilética que sugiere una liberación de limitaciones estructurales. Segundo, el antepasado común ya era una célula asombrosamente compleja. Mediante la comparación de características comunes a cada uno de los supergrupos, podemos reconstruir las propiedades probables del antepasado común. Cualquier característica presente esencialmente en todas las especies de todos los supergrupos se heredó presumiblemente de aquel antepasado común, mientras que cabe presumir que cualesquiera rasgos que sólo estén presentes en uno o dos grupos se adquirieron posteriormente, y sólo en aquel grupo. Los cloroplastos son un buen ejemplo de esto último: se encuentran sólo en las plantas y los cromalveolados, como resultado de endosimbiosis bien conocidas. No formaban parte del antepasado eucariota común.
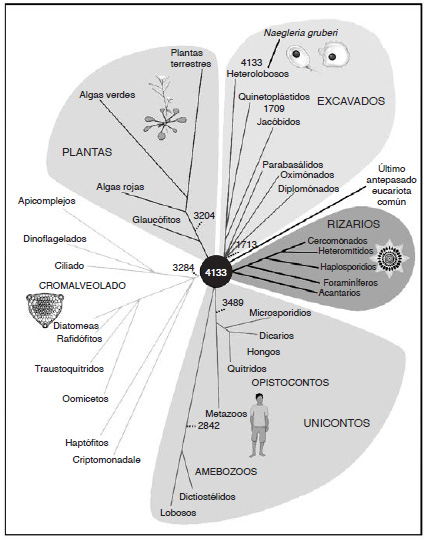
FIG. 5. Los «supergrupos» de eucariotas. Un árbol de los eucariotas, basado en miles de genes compartidos, que muestra los cinco «supergrupos», tal como lo diseñó Eugene Koonin en 2010. Los números hacen referencia al número de genes compartidos por cada uno de estos supergrupos con el LECA (el último antepasado eucariota común). Cada grupo ha perdido o ganado de forma independiente otros muchos genes. Aquí la mayor variación se da entre los protistas unicelulares; los animales se encuentran dentro de los metazoos (en la parte inferior). Adviértase que hay mucha más variación dentro de cada supergrupo que entre los antepasados de dichos grupos, lo que sugiere una radiación explosiva temprana. Me gusta el agujero negro simbólico en el centro: el LECA ya había adquirido por evolución todos los rasgos eucariotas comunes, pero la filogenia da pocas indicaciones acerca de cómo surgieron dichos rasgos, de las bacterias o de los arqueos: un agujero negro evolutivo.
Así pues, ¿qué nos dice la filogenia que formaba parte del antepasado común? Asombrosamente, casi todo lo demás. Permítaseme repasar unos pocos casos. Sabemos que el antepasado común tenía un núcleo, en el que almacenaba su DNA. El núcleo tiene una gran cantidad de estructura compleja que de nuevo se conserva en todos los eucariotas. Está encerrado por una doble membrana, o más bien una serie de sacos aplanados que parecen una doble membrana pero que en realidad se continúan en otras membranas celulares. La membrana nuclear está tachonada con complejos poros de proteína y revestida por una matriz elástica; y dentro del núcleo, otras estructuras, como el nucléolo, se conservan asimismo en todos los eucariotas. Vale la pena resaltar que docenas de proteínas básicas en estos complejos se conservan en todos los supergrupos, como son las histonas que rodean el DNA. Todos los eucariotas poseen cromosomas rectos, rematados por «telómeros», que evitan que los extremos se deshilachen como los de los cordones de los zapatos. Los eucariotas tienen «genes en fragmentos», en los que secciones cortas de proteínas que codifican DNA están entremezcladas con largas regiones no codificadoras, llamadas intrones. Estos intrones son empalmados antes de ser incorporados a proteínas, utilizando una maquinaria común a todos los eucariotas. Incluso se suele conservar la posición de los intrones, con inserciones que se encuentran en la misma posición del mismo gen en todos los eucariotas.
Fuera del núcleo, se repite el mismo tipo de situación. Excepto los arquezoos más simples (que resultan estar muy repartidos en los cinco supergrupos, con lo que se muestra de nuevo su pérdida independiente de la complejidad primitiva), todos los eucariotas comparten esencialmente la misma maquinaria celular. Todos poseen complejas estructuras membranosas internas, como el retículo endoplasmático y el aparato de Golgi, que están especializados en empaquetar y exportar proteínas. Todos poseen un citoesqueleto interno dinámico, capaz de remodelarse en todas las formas y necesidades. Todos tienen proteínas motrices que trasladan objetos de un lado a otro siguiendo pistas citoesqueléticas a través de la célula. Todos tienen mitocondrias, lisosomas, peroxisomas, la maquinaria de importación y exportación, y sistemas comunes de señalización. La lista sigue. Todos los eucariotas se dividen mediante mitosis, en la que los cromosomas se separan sobre un huso microtubular, utilizando un conjunto de enzimas común. Todos son sexuales, con un ciclo biológico que implica meiosis (división reductiva) para formar gametos como los espermatozoides y los óvulos, seguida por la fusión de dichos gametos. Los pocos eucariotas que pierden su sexualidad tienden a extinguirse rápidamente (en este caso, rápidamente significa unos pocos millones de años).
Gran parte de esto lo hemos sabido desde hace mucho tiempo a partir de la estructura microscópica de las células, pero la nueva era de la filogenómica ilumina de manera vigorosa dos aspectos. Primero, las semejanzas estructurales no son parecidos superficiales, aspectos que quedan bien para engañar, sino que están escritos en las secuencias detalladas de los genes, en millones y miles de millones de letras de DNA, y esto nos permite computar su abolengo, con una precisión sin precedentes, como un árbol que se ramifica. Segundo, la aparición de la secuenciación génica de alto rendimiento significa que el muestreo del mundo natural ya no se basa en laboriosos intentos de cultivar células o de preparar secciones microscópicas, sino que es tan rápido y fiable como un secuenciador por fuerza bruta. Hemos descubierto varios grupos nuevos e insospechados, entre ellos eucariotas extremófilos capaces de habérselas con altas concentraciones de metales tóxicos o con temperaturas elevadas, y células diminutas pero perfectamente formadas conocidas como picoeucariotas, tan pequeñas como bacterias pero que presentan un núcleo también reducido y mitocondrias enanas. Todo esto significa que tenemos una idea mucho más clara de la diversidad de los eucariotas. Todos estos eucariotas nuevos encajan de manera cómoda dentro de los cinco supergrupos establecidos: no abren nuevos panoramas filogenéticos. El hecho magnífico que surge de esta enorme diversidad es lo condenadamente similares que son las células eucariotas. No encontramos todo tipo de formas intermedias y de variantes no emparentadas. La predicción de la teoría de la endosimbiosis serial, según la cual sí las encontraríamos, está equivocada.
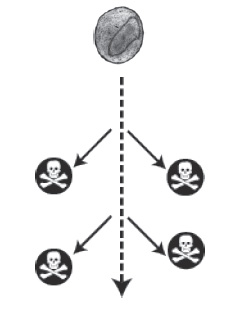
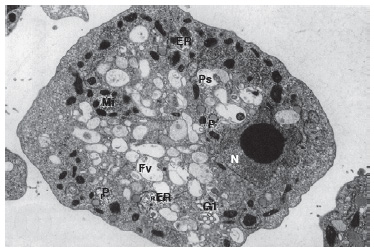
FIG. 6. El agujero negro en el meollo de la biología. La célula de la parte inferior es Naegleria, que se cree que es parecida en tamaño y complejidad al antepasado común de todos los eucariotas. Posee un núcleo (N), retículo endoplasmático (ER), complejo de Golgi (Gl), mitocondrias (Mi), vacuola alimentaria (Fv), fagosomas (Ps) y peroxisomas (P). En la parte superior hay una bacteria relativamente compleja, Planctomycetes, que se muestra aproximadamente a escala. No sugiero que los eucariotas derivaron de Planctomycetes (ciertamente, no lo hicieron), sino que simplemente muestro la escala de la brecha entre una bacteria relativamente compleja y un eucariota unicelular representativo. No hay estadios evolutivos intermedios supervivientes que nos cuenten la historia (lo que se indica mediante las calaveras y los huesos cruzados).
Esto plantea un problema diferente. El asombroso éxito de la filogenia y la aproximación informacional a la biología pueden cegarnos fácilmente ante sus limitaciones. El problema aquí es el equivalente de un «horizonte de eventos» filogenético en el origen de los eucariotas. Todos estos genomas conducen al último antepasado común de los eucariotas, que lo tenía más o menos todo. Pero ¿de dónde procedían estas partes? El antepasado común de los eucariotas bien podría haber saltado, completamente formado, como Atenea de la cabeza de Zeus. Tenemos poca información acerca de los rasgos que surgieron antes del antepasado común… esencialmente todos ellos. ¿Cómo y por qué evolucionó el núcleo? ¿Y qué hay del sexo? ¿Por qué prácticamente todos los eucariotas tienen dos sexos? ¿De dónde proceden las extravagantes membranas internas? ¿Cómo se hizo tan dinámico y flexible el citoesqueleto? ¿Por qué la división sexual celular («meiosis») divide el número de cromosomas después de haberlo duplicado? ¿Por qué envejecemos, contraemos cáncer y morimos? A pesar de todo su ingenio, la filogenia puede decirnos poca cosa acerca de estas preguntas fundamentales de la biología. Casi todos los genes implicados (que codifican las llamadas «proteínas de rúbrica» de los eucariotas) no se encuentran en los procariotas. Y, a la inversa, las bacterias no muestran prácticamente tendencia alguna a producir por evolución ninguno de estos rasgos complejos de los eucariotas. No existen formas evolutivas intermedias conocidas entre el estado morfológicamente simple de todos los procariotas y el antepasado común de los eucariotas, inquietantemente complejo (figura 6). Todos estos atributos de la vida compleja surgieron en un vacío filogenético, un agujero negro en el meollo de la biología.
Los pasos que faltan hasta la complejidad
La teoría evolutiva hace una predicción simple. Las características complejas surgen a través de una serie de pequeños pasos, y cada nuevo paso ofrece una pequeña ventaja sobre el anterior. La selección de las características mejor adaptadas significa la pérdida de los rasgos menos bien adaptados, de modo que la selección elimina continuamente formas intermedias. A lo largo del tiempo, las características tenderán a escalar las cumbres de un paisaje adaptativo, de modo que vemos la aparente perfección de los ojos, pero no los pasos intermedios menos perfectos en route hacia su evolución. En El origen de las especies, Darwin planteó el hecho de que la selección natural predice realmente que las formas intermedias han de perderse. En este contexto, no es terriblemente sorprendente que no existan formas intermedias supervivientes entre las bacterias y los eucariotas. Lo que es más sorprendente, sin embargo, es que los mismos rasgos no vayan surgiendo, una y otra vez… como los ojos.
No vemos los pasos históricos en la evolución de los ojos, pero sí que vemos un espectro ecológico. A partir de un punto rudimentario sensible a la luz en algún animal primitivo parecido a un gusano, los ojos han surgido de manera independiente en multitud de ocasiones. Esto es exactamente lo que predice la selección natural. Cada pequeño paso ofrece una pequeña ventaja en un ambiente concreto, y la ventaja precisa depende del ambiente preciso. Tipos de ojos morfológicamente distintos han surgido por evolución en ambientes diferentes, tan divergentes como los ojos compuestos de las moscas y los ojos en espejo de las vieiras, o tan convergentes como los ojos en cámara que son tan parecidos en los humanos y los pulpos. Todas las formas intermedias imaginables, desde agujeritos hasta lentes ajustables, se encuentran en una especie u otra. Hemos visto incluso ojos en miniatura, completos con una «lente» y una «retina», en algunos protistas unicelulares. En resumen, la teoría evolutiva predice que debe haber orígenes múltiples (polifiléticos) de aquellas características en las que cada pequeño paso ofrece una pequeña ventaja sobre el paso anterior. En teoría, esto se aplica a todas las características, y en efecto es lo que vemos de manera general. De modo que el vuelo batido surgió al menos en seis ocasiones diferentes, en murciélagos, aves, pterosaurios y varios insectos; la pluricelularidad unas 30 veces, tal como se indicó anteriormente; diferentes formas de endotermia (sangre caliente) en varios grupos, entre ellos las aves y los mamíferos, pero también en algunos peces, insectos y plantas;13 e incluso la percepción consciente parece haber surgido de manera más o menos independiente en aves y mamíferos. Al igual que con los ojos, vemos una multitud de formas diferentes que reflejan los ambientes diferentes en los que surgieron. Existen, ciertamente, limitaciones físicas, pero no son lo bastante fuertes para descartar orígenes múltiples.
Así pues, ¿qué ocurre con el sexo, o el núcleo, o la fagocitosis? Cabría aplicar el mismo razonamiento. Si cada uno de estos rasgos surgió mediante selección natural (y ello es indudable) y todos los pasos adaptativos ofrecieron alguna pequeña ventaja (y ello es indudable), entonces tendríamos que ver orígenes múltiples de rasgos de los eucariotas en las bacterias. Pero no los vemos. Esto es casi un «escándalo» evolutivo. En las bacterias no vemos más que los inicios de rasgos eucariotas. Tomemos el sexo, por ejemplo. Hay quien puede aducir que las bacterias practican una forma de conjugación equivalente al sexo, transfiriendo DNA de una bacteria a otra mediante transferencia génica «lateral». Las bacterias tienen toda la maquinaria necesaria para recombinar el DNA, lo que les permite forjar nuevos y variados cromosomas, que por lo general se considera que es la ventaja del sexo. Pero las diferencias son enormes. El sexo implica la fusión de dos gametos, cada uno de ellos con la mitad de la dotación normal de genes, seguida de recombinación recíproca en todo el genoma. La transferencia génica lateral no es en este sentido recíproca ni sistemática, sino fragmentada. En efecto, los eucariotas practican el «sexo total», y las bacterias una forma tibia y pálida. Es claro que ha de existir alguna ventaja para que los eucariotas se dediquen al sexo total; pero si así fuera, cabría esperar que al menos algunos tipos de bacterias hicieran algo similar, incluso si en su detalle los mecanismos fueran diferentes. Hasta donde sabemos, ninguna lo hizo nunca. Lo mismo es válido para el núcleo y la fagocitosis, y más o menos para todas las características propias de los eucariotas. Los primeros pasos no son el problema. Vemos algunas bacterias con membranas internas plegadas, otras sin pared celular y un citoesqueleto modestamente dinámico, y aun otras con cromosomas rectos, o con copias múltiples de su genoma, o con un tamaño celular gigantesco: todos los inicios de la complejidad eucariota. Pero las bacterias siempre se detienen mucho antes de la complejidad barroca de los eucariotas, y raramente, si acaso lo hacen, combinan rasgos complejos múltiples en la misma célula.
La explicación más fácil para las profundas diferencias entre bacterias y eucariotas es la competencia. Según este razonamiento, una vez hubieron surgido por evolución los primeros eucariotas verdaderos fueron tan competitivos que dominaron el nicho de la complejidad morfológica. Ninguna otra cosa podía competir con ellos. A cualquier bacteria que «intentara» invadir este nicho eucariota se le prestaba poca atención por parte de las células sofisticadas que ya vivían allí. Para usar la expresión común, perdieron en la competencia hasta extinguirse. Todos estamos familiarizados con las extinciones en masa de los dinosaurios y otros grandes animales y plantas, de modo que esta explicación parece perfectamente razonable. Los pequeños y peludos antepasados de los mamíferos modernos fueron mantenidos a raya por los dinosaurios durante millones de años, y sólo radiaron en los grupos modernos después de la desaparición de los dinosaurios. Pero existen buenas razones para poner en cuestión esta idea razonable, pero engañosa. Los microbios no son equivalentes a los animales grandes: los tamaños de sus poblaciones son enormemente mayores, y se transmiten genes útiles (como los correspondientes a la resistencia a los antibióticos) mediante transferencia génica lateral, lo que los hace mucho menos vulnerables a la extinción. No existe indicación alguna de ninguna extinción microbiana, incluso después del Gran Evento de Oxidación. No existe indicio alguno del «holocausto del oxígeno», que supuestamente aniquiló a la mayoría de las células anaerobias: no hay pruebas procedentes de la filogenia ni de la geoquímica de que tal extinción tuviera nunca lugar. Por el contrario, los anaerobios prosperaron.
Más importante todavía: de hecho, hay pruebas muy consistentes de que las formas intermedias no se extinguieron debido a la competencia por parte de los eucariotas más complejos. Todavía existen. Ya nos encontramos con ellos: los «arquezoos», el gran grupo de eucariotas primitivos que antaño fueron considerados erróneamente un eslabón perdido. No son verdaderas formas evolutivas intermedias, sino que son formas ecológicas intermedias reales. Ocupan el mismo nicho. Una forma evolutiva intermedia es un eslabón perdido: un pez con patas, como Tiktaalik, o un dinosaurio con plumas y alas, como Archaeopteryx. Una forma ecológica intermedia no es un verdadero eslabón perdido pero demuestra que un nicho ecológico, una forma de vida determinados, son viables. Una ardilla voladora no está estrechamente emparentada con otros vertebrados voladores como los murciélagos o las aves, pero demuestra que el vuelo planeado entre árboles es posible sin alas completamente desarrolladas. Ello significa que no es una pura fantasía sugerir que el vuelo batido pudo haberse iniciado de esta manera. Y esta es la importancia real de los arquezoos: son formas ecológicas intermedias, que demuestran que una determinada forma de vida es viable.
Mencioné anteriormente que hay un millar o más de especies diferentes de arquezoos. Estas células son eucariotas bona fide, que se adaptaron a este nicho «intermedio» haciéndose más simples, no bacterias que se hicieron algo más complejas. Permítaseme insistir en este punto. El nicho es viable. Ha sido invadido en numerosas ocasiones por células morfológicamente sencillas, que medran allí. Estas células sencillas no se extinguieron por la competencia de eucariotas más complejos que ya existían y ocupaban el mismo nicho. Más bien lo contrario: prosperaron precisamente porque se hicieron más simples. En términos estadísticos, si todo lo demás no varía, la probabilidad de que únicamente eucariotas sencillos (en lugar de bacterias complejas) invadan este nicho en 1.000 ocasiones distintas es del orden de uno en 10300 en contra, un número que podía haber sido conjurado por la Fuerza de Infinita Improbabilidad de Zaphod Beeblebrox.14 Incluso si los arquezoos surgieron de manera independiente en unas más conservadoras 20 ocasiones distintas (cada vez radiando para producir un número grande de especies hijas), la probabilidad sigue siendo de una en un millón en contra. O bien esto fue una chiripa de extravagantes proporciones, o todo lo demás variaba. La explicación más plausible es que había algo en la estructura de los eucariotas que facilitó su invasión de este nicho intermedio, y al revés, algo acerca de la estructura de las bacterias que impidió su evolución de una mayor complejidad morfológica.
Esto no parece particularmente radical. En realidad, pega con todo lo demás que conocemos. A lo largo de este capítulo he hablado de bacterias, pero como señalamos en la Introducción, de hecho existen dos grandes grupos o dominios de células que carecen de núcleo, y por tanto son designados como «procariotas» (literalmente, «antes del núcleo»). Son las bacterias y los «arqueos», que no hay que confundir con los arquezoos, las células eucariotas sencillas de las que hemos estado hablando. Mientras que sólo puedo pedir disculpas por la confusión de terminología científica, que a veces parece ser elaborada por alquimistas que ansían no ser comprendidos, recuerde por favor el lector que los arqueos y las bacterias son procariotas, que carecen de núcleo, mientras que los arquezoos son eucariotas primitivos, que poseen un núcleo. De hecho, a los arqueos todavía se les llama arquebacterias, o «bacterias antiguas», en oposición a las eubacterias, o «bacterias verdaderas», de manera que a ambos grupos se los puede denominar legítimamente bacterias. En aras de la simplicidad, continuaré empleando el término bacterias de manera laxa para referirme a ambos grupos, excepto cuando necesite especificar diferencias críticas entre los dos dominios.15
El punto crucial es que estos dos dominios, las bacterias y los arqueos, son extremadamente diferentes en su genética y en su bioquímica, pero casi indistinguibles por su morfología. Ambos tipos son células pequeñas y sencillas que carecen de núcleo y de todos los demás rasgos eucariotas que definen la vida compleja. El hecho de que ambos grupos no consiguieran desarrollar una morfología compleja, a pesar de su diversidad genética y de su ingeniosidad bioquímica extraordinarias, hace que parezca como si una limitación física intrínseca impida la evolución de la complejidad en los procariotas, una limitación que de alguna manera desapareció en la evolución de los eucariotas. En el capítulo 5 argumentaré que esta limitación fue liberada por un acontecimiento extraordinario: la endosimbiosis singular entre dos procariotas de la que ya discutimos en la Introducción. Sin embargo, por ahora señalemos simplemente que algún tipo de limitación estructural tuvo que haber actuado igualmente en ambos grandes dominios de los procariotas, las bacterias y los arqueos, lo que obligó a ambos grupos a permanecer simples en su morfología a la largo de unos incomprensibles 4.000 millones de años. Sólo los eucariotas exploraron el ámbito de la complejidad, y lo hicieron mediante una radiación monofilética explosiva que implica una liberación de lo que fuera que estas limitaciones estructurales pudieran haber sido. Parece que esto ocurrió una sola vez: todos los eucariotas están emparentados.
La pregunta equivocada
Esta, pues, es nuestra breve historia de la vida a través de ojos nuevos. He aquí un resumen rápido. La Tierra primitiva no era drásticamente diferente de nuestro propio mundo: era un mundo acuático, con un clima moderado, dominado por gases volcánicos como dióxido de carbono y nitrógeno. Aunque nuestro planeta primitivo carecía de oxígeno, no era rico en gases propicios para la química orgánica: hidrógeno, metano y amoníaco. Esto descarta ideas antiguas y gastadas de una sopa primordial; pero la vida empezó tan pronto como pudo, hace quizá cuatro mil millones de años. En un sentido literal, alguna otra cosa impulsaba la aparición de la vida; ya llegaremos a ello. Las bacterias pronto dominaron, colonizando cada centímetro, cada nicho metabólico, remodelando el globo a lo largo de dos mil millones de años, depositando rocas y minerales a una escala colosal, y transformando los océanos, la atmósfera y los continentes. Alteraron drásticamente el clima en Tierras bola de nieve globales; oxidaron el mundo, llenando océanos y aire con oxígeno reactivo. Pero a lo largo de toda esta inmensa duración, ni las bacterias ni los arqueos se convirtieron en otra cosa: permanecieron tozudamente simples en su estructura y modo de vida. Durante unos eternos cuatro mil millones de años, pasando por extremos de cambio ambiental y ecológico, las bacterias cambiaron sus genes y su bioquímica, pero no cambiaron nunca su forma. Nunca dieron origen a seres vivos más complejos, del tipo que podríamos esperar detectar en otro planeta, extraterrestres inteligentes… excepto por una única vez.
En una única ocasión, aquí en la Tierra, las bacterias dieron origen a los eucariotas. No hay nada en el registro fósil, ni en la filogenia, que sugiera que la vida compleja surgió realmente de manera repetida, pero que sólo un grupo, los eucariotas modernos y familiares, sobrevivió. Por el contrario, la radiación monofilética de los eucariotas sugiere que su origen único fue dictado por restricciones físicas innatas que tuvieron poco o nada que ver con trastornos ambientales como el Gran Evento de Oxidación. En la parte III del libro veremos cuáles pudieron haber sido estas restricciones. Por ahora, digamos simplemente que cualquier relato adecuado ha de explicar por qué la evolución de la vida compleja tuvo lugar únicamente una vez: nuestra explicación ha de ser lo bastante persuasiva para ser creíble, pero no tan persuasiva que nos quedemos preguntándonos por qué no ocurrió en muchas ocasiones. Cualquier intento de explicar un acontecimiento singular tendrá siempre el aspecto de una chiripa. ¿Cómo podemos demostrarlo de una u otra manera? Puede que el acontecimiento mismo no dé mucho de sí, pero puede haber pistas escondidas en sus repercusiones, una pistola humeante que proporcione algún indicio de lo que ocurrió. Una vez se libraron de sus grilletes bacterianos, los eucariotas se hicieron enormemente complejos y diversos en su morfología. Pero no aumentaron su complejidad de una manera que fuera obviamente predecible: aparecieron con toda una serie de rasgos, desde el sexo y el envejecimiento a la especiación, ninguno de los cuales se había visto nunca en las bacterias o los arqueos. Los eucariotas más antiguos acumularon todos estos rasgos singulares en un antepasado común sin parangón. No existen formas intermedias evolutivas conocidas entre la simplicidad morfológica de las bacterias y aquel antepasado común eucariota enormemente complejo que puedan contarnos la historia. Todo esto se suma a una posibilidad apasionante: ¡las mayores preguntas en biología siguen sin resolverse! ¿Acaso existe en estas características algún patrón que pueda dar una indicación de cómo evolucionaron? Así lo creo.
Este enigma está relacionado con la pregunta que planteamos al principio de este capítulo. ¿Qué parte de la historia y de las propiedades de la vida puede predecirse a partir de primeros principios? Sugerí que la vida está limitada de maneras que no pueden ser fácilmente interpretadas en términos de genomas, historia o ambiente. Si consideramos la vida únicamente en términos de información, mi opinión era que no podríamos predecir nada de su historia inescrutable. ¿Por qué empezó tan pronto la vida? ¿Por qué se estancó en estructura morfológica durante miles de millones de años? ¿Por qué bacterias y arqueos no resultaron afectados por trastornos ambientales y ecológicos que ocurrieron a una escala global? ¿Por qué toda la vida compleja es monofilética, y surgió sólo una vez en 4.000 millones de años? ¿Por qué los procariotas no dan origen continuamente, o incluso ocasionalmente, a células y organismos con mayor complejidad? ¿Por qué rasgos individuales propios de los eucariotas, como el sexo, el núcleo y la fagocitosis, no surgieron en las bacterias o los arqueos? ¿Por qué los eucariotas acumularon todos estos rasgos?
Si la vida trata de información, estos son misterios profundos. No creo que este relato pudiera haberse pronosticado, predicho como ciencia, únicamente sobre la base de la información. Las propiedades extravagantes de la vida tendrían que adscribirse a las contingencias de la historia, a los golpes y dardos de la insultante fortuna.16 No tendríamos posibilidad de predecir las propiedades de la vida en otros planetas. Pero el DNA, el seductor código que parece prometer todas las respuestas, nos ha hecho olvidar el otro principio fundamental de Schrödinger: que la vida resiste la entropía, la tendencia a la descomposición. En una nota de pie de página a What is Life?, Schrödinger advertía que si hubiera escrito para una audiencia de físicos hubiera planteado su argumentación no en términos de entropía, sino de energía libre. El término «libre» tiene un significado específico, que consideraremos en el capítulo siguiente; por ahora baste decir que la energía es precisamente lo que faltaba en este capítulo, y de hecho en el libro de Schrödinger. Su famoso título hacía la pregunta totalmente equivocada. Si se añade la energía, la pregunta es mucho más contundente: What is Living?17 Pero hay que perdonar a Schrödinger. No podía haberlo sabido. Cuando escribía, nadie sabía demasiado acerca de la moneda biológica de la energía. Ahora sabemos cómo funciona todo en un detalle exquisito, hasta el mismo nivel de los átomos. Los mecanismos detallados de la obtención de energía resultan conservarse tan universalmente en toda la vida como el propio código genético, y dichos mecanismos ejercen limitaciones fundamentales sobre las células. Pero no tenemos ni idea de cómo evolucionaron, ni de qué manera la energía biológica limitó el relato de la vida. De esto es de lo que trata este libro.