capítulo 2
ANATOMÍA DEL DESARROLLO
Dra. Marcia Gaete Carrasco
Gametogénesis
La gametogénesis es un proceso que consta de cuatro fases: Origen y migración de las células germinales hacia las gónadas, incremento del número de células germinales, reducción cromosómica y maduración de los gametos.
Origen y migración de las células germinales hacia las gónadas
Las células que formarán los gametos en ambos sexos derivan de las llamadas células germinales primordiales. Ellas se originan en una hoja del embrión llamada epiblasto o ectodermo primitivo, durante la segunda semana de desarrollo. Luego las células germinales primordiales se trasladan a la pared de una estructura extraembrionaria llamada saco vitelino. Entre la cuarta y quinta semana de vida intrauterina migran por el mesenterio primitivo hacia la pared posterior del abdomen, en dirección a la futura región gonadal, donde estimulan la formación de las crestas genitales. Allí se desarrollarán las gónadas.
Incremento del número de células germinales
En las crestas genitales las células germinales sufren sucesivas mitosis antes de entrar en el proceso de gametogénesis, que las convierte en gametos femeninos o masculinos maduros: ovocitos maduros o espermatozoides. En el hombre, recién en la pubertad, las células germinales comienzan a diferenciarse en espermatogonias que pueden entrar en meiosis. En la mujer, las células germinales se diferencian a ovogonias al quinto mes de desarrollo fetal, donde comienzan la meiosis, pero quedan en reposo hasta la pubertad donde, en grupos, retoman la meiosis con cada ciclo ovárico.
Reducción cromosómica
Desde las células germinales se diferencian las ovogonias o espermatogonias. La gametogénesis se inicia cuando las ovogonias o espermatogonias ingresan a la meiosis, recibiendo el nombre de ovocitos y espermatocitos primarios. Como se mencionó anteriormente, en los hombres, este proceso empieza en la pubertad y es cíclico. En la mujer la población de ovocitos primarios se establece al nacer y la meiosis es frenada para ser reactivada en la pubertad. Los cambios cromosómicos que ocurren en la gametogénesis involucran: la reducción del número de cromosomas mediante la meiosis generándose células haploides con 22 autosomas y 1 cromosoma sexual. La meiosis es un tipo de división celular exclusiva de los gametos y permite el crossing over o entrecruzamiento, proceso en el que se recombinan los genes aumentando la variabilidad entre individuos. En el caso del hombre, la meiosis permite también generar espermatozoides con un cromosoma sexual X o Y, lo que determina el sexo del cigoto en la fecundación. El gameto femenino siempre aporta un cromosoma X, por lo que si el espermatozoide porta un cromosoma X el cigoto será femenino (XY) y si porta un cromosoma Y será masculino (XY).
La meiosis presenta dos divisiones de maduración: la primera división es reduccional porque desde una célula diploide, sea ovocito o espermatocito primario, se generan dos células haploides: dos espermatocitos secundarios en el caso del hombre y un ovocito secundario y un cuerpo polar en el caso de la mujer. De la segunda división, que es ecuacional, a partir de cada ovocito o espermatocito secundario se generan dos células haploides: dos espermátides en el caso del hombre o un ovocito maduro y un segundo cuerpo polar en el caso de la mujer.
Maduración de los gametos
Los gametos en su maduración desarrollan especializaciones que los habilitan para participar en la fecundación. Los cambios de maduración que ocurren durante la gametogénesis masculina para formar los espermatozoides son: la condensación del núcleo, la eliminación de gran parte del citoplasma, formación del acrosoma y diferenciación de una célula alargada, con capacidad de moverse, con cabeza, cuello, pieza intermedia y cola. El acrosoma es una vesícula ubicada en el extremo apical de la célula, delante del núcleo, que contiene enzimas capaces de digerir la zona pelúcida, una densa matriz que rodea al ovocito, permitiendo la fecundación.
Durante la gametogénesis femenina las especializaciones celulares determinan la formación de un óvulo que se caracteriza por ser inmóvil, poseer un tamaño enorme (150 µm) –comparado con el espermatozoide que mide 3 µm–, presentar una gran cantidad de citoplasma, estar rodeado por una capa glicoproteica (zona pelúcida), que le confiere la especificidad para la fecundación, y una capa de células foliculares: la corona radiada.
Gametogénesis masculina
La gametogénesis masculina o espermatogénesis es el proceso de división y diferenciación celular meiótico que conduce a la formación de espermatozoides. Esta comienza en la pubertad y continúa durante toda la vida. Las células germinales se multiplican por divisiones mitóticas seguidas por divisiones meióticas que produce el estado haploide, esto va acompañado con una migración de las células desde la pared del túbulo seminífero al lumen.
Así, las espermatogonias se localizan en la zona más externa del túbulo seminífero, próximas a la membrana basal, entre las células de Sertoli. Las espermatogonias A, localizadas más externamente, son las células madre. Las espermatogonias B, originadas por mitosis de las células A, se dividen para formar para formar espermatocitos primarios. Cada espermatocito primario se divide para formar dos espermatocitos secundarios, cambiando el número de cromosomas a haploides. Luego los espermatocitos secundarios forman dos espermátidas cada uno. Durante la espermatogénesis el núcleo y el citoplasma de la espermátida pasa por cambios morfológicos para formar el espermatozoide (refiérase a Maduración de los gametos, descrita previamente). Al final de la gametogénesis el espermatozoide se pone en contacto con el lumen del túbulo seminífero y se desprende como célula independiente (Figura 2-1).
Gametogénesis femenina
Cuando las germinales primordiales alcanzan la región de la gónada femenina comienzan a diferenciarse en ovogonias. Las ovogonias se agrupan en la corteza del ovario, hacen mitosis y al cuarto mes se reconocen cúmulos de células hermanas que provienen de una sola ovogonia rodeadas por una monocapa de células epiteliales planas, derivadas del epitelio superficial del ovario. Algunas de estas ovogonias entran en meiosis formando ovocitos primarios. Durante el quinto mes de desarrollo, los ovocitos primarios se rodean cada uno por las células epiteliales planas, llamadas ahora foliculares, para constituir el llamado folículo primordial. Al quinto mes de vida intrauterina, ovogonios y ovocitos alcanzan un máximo de 7 millones y luego degeneran en un proceso fisiológico llamado atresia, de modo tal que al momento del nacimiento, el número de ovocitos es alrededor de 800.000 y cuando se alcanza la pubertad el número se reduce a 40.000.
Cerca del nacimiento, cada uno de estos ovocitos entran en una etapa de reposo durante la fase de diploteno de la primera división meiótica. Estas células se mantendrán en un estado latente durante muchos años, hasta la pubertad, donde retoman su meiosis cuando comienzan los ciclos ováricos.
Ciclo ovárico
A partir de la pubertad, la mujer presenta ciclos ováricos. Estos ciclos preparan el sistema reproductor para el embarazo e involucran al hipotálamo a través de la hormona liberadora de gonadotrofinas (GRH, por su sigla en inglés); a la hipófifis, estimulada por la GRH para que secrete gonadotrofinas: hormona foliculoestimulante (FSH, por su sigla en inglés) y hormona luteinizante (LH, por su sigla en inglés), que actúan sobre los ovarios, útero, vagina, tubas uterinas y glándulas mamarias.
Al iniciarse el ciclo (entre los días 1 al 6 del ciclo menstrual), la FSH induce el desarrollo de 5 a 10 folículos ováricos, constituidos por el ovocito rodeado por células foliculares, pero solo uno llega a madurar. Los folículos que no se desarrollan involucionan y forman los llamados cuerpos atrésicos.
Las células foliculares forman la zona pelúcida, una capa glicoproteica que las separa del ovocito. Si bien esta es una membrana continua, contiene prolongaciones de las células foliculares que se ponen en contacto con el ovocito. Las células que rodean a la zona pelúcida y al ovocito primario constituyen el grupo de células de la granulosa. La primera capa de la granulosa que toma ontacto con el ovocito es la corona radiada. Las células de la granulosa, la zona pelúcida y el ovocito primario conforman así el denominado folículo primario (Figuras 2-2a y b).
Al continuar la estimulación de la hormona foliculoestimulante las células de la granulosa, que van aumentando en número, comienzan a tener espacios entre sí y determinan la conformación de una cavidad denominada antro folicular, conformando un folículo secundario. Finalmente, el folículo secundario constituye una gran cavidad o antro que se va haciendo cada vez más amplia y determina que este grupo de células de la granulosa, con su zona pelúcida y con su ovocito primario, sean desplazadas a un extremo del folículo. Las células de la granulosa, a su vez, constituyen el denominado cúmulo oóforo. Un folículo que presenta estas características, o sea, que tiene un cúmulo oóforo, posee células granulosas en la periferia y un tejido conjuntivo ovárico especializado, llamado teca, se denomina folículo terciario o de Graaf, y es el que será ovulado.
Los días previos a la ovulación (días 12 a 13 del ciclo menstrual), el folículo de Graaf aumenta de tamaño, llegando a medir 15 mm. Este crecimiento es estimulado por las hormonas FSH y LH. En esta etapa el ovocito primario completa su primera división meiótica, generándose así el ovocito secundario, que tiene la mayor parte del citoplasma, y el primer cuerpo polar, célula con poco citoplasma.
El folículo de Graaf se ubica en la superficie del ovario y comienza a presionar, formando un estigma. Finalmente el ovocito es expulsado junto a su cúmulo oóforo al día 14 del ciclo aproximadamente. En este momento el ovocito secundario empieza la segunda división meiótica.
Desde la superficie del ovario, el ovocito es tomado por la tuba uterina y, mediante movimientos peristálticos de la tuba uterina y movimientos ciliares de la mucosa, es transportado hacia el lumen de la tuba uterina.
El resto de folículo, que no es secretado, queda en el estroma ovárico constituyendo de esta forma el denominado cuerpo lúteo o amarillo; que secreta progesterona hasta aproximadamente el noveno día después de la ovulación, permitiendo que el endometrio entre en la etapa progestacional o secretora, aumentando la vascularización y secreción de las glándulas mucosas. Si ocurre embarazo el cuerpo lúteo crece, secretando grandes cantidades de progesterona y constituyendo entonces el cuerpo gravídico, que puede verse como una gran mancha amarilla que al tercer mes llega a ocupar un tercio del ovario; posteriormente involuciona y la placenta asume el rol de secretora de progesterona. Si el cuerpo gravídico se extirpara antes del cuarto mes de desarrollo fetal, produce aborto.
Primera semana del desarrollo: fecundación, segmentación
e implantación
Fecundación
La fecundación es el proceso mediante el cual se fusionan el gameto masculino o espermatozoide y el gameto femenino u ovocito, originando un huevo o cigoto. Ocurre generalmente en la región ampular de la tuba. Los espermatozoides pueden mantenerse viables en el tracto femenino hasta por cinco a siete días, en cambio el ovocito luego de la ovulación permanece viable por 24 horas. Aunque cerca de 300 millones de espermatozoides son depositados en el tracto genital femenino aproximadamente unos 500 llegan a las proximidades del ovocito.
Para que ocurra la fecundación es necesario que los espermatozoides experimenten el proceso de capacitación y de reacción acrosómica.
La capacitación ocurre en el tracto genital femenino y dura 7 horas. Durante la capacitación los espermatozoides liberan glucoproteínas presentes en la superficie del acrosoma y comienzan a oscilar activamente para atravesar las células de la corona radiante. Los gametos capacitados, al tomar contacto con la zona pelúcida, experimentan la reacción acrosómica, liberando enzimas presentes en el acrosoma que le permitirán atravesarla y llegar a la membrana celular del ovocito maduro. Una vez que el espermatozoide atraviesa la zona pelúcida y toca la membrana celular del ovocito, la zona pelúcida se hace impermeable a otros espermatozoides, es decir, hay un bloqueo de la poliespermia. Existe un bloqueo rápido de la poliespermia que es mediado por cambios en el potencial de acción del óvulo que se hace más positivo, de -70 a +10 mV. El bloqueo lento de la poliespermia es mediado por la reacción de zona, proceso que, gatillado por la liberación de enzimas lisosomales de los gránulos corticales del óvulo, induce un cambio químico en la zona pelúcida, la que se hace menos adherente a los espermatozoides. Muchos de los espermatozoides capacitados atraviesan la corona radiante y algunos se sumergen parcialmente en la zona pelúcida, pero únicamente uno de ellos atravesará completamente la zona pelúcida y la membrana plasmática del ovocito (Figuras 2-3a, b y c).
En este momento las membranas celulares de los gametos se fusionan, permitiendo que ingrese la cabeza y la cola del espermatozoide al citoplasma del ovocito. La fusión ocurre entre la membrana del ovocito y la región posterior de la cabeza del espermatozoide, debido a que la membrana plasmática de la punta de la cabeza desaparece con la reacción acrosómica. Cuando este proceso ocurre, el ovocito completa su segunda división meiótica, formando el segundo cuerpo polar. El ovocito así pasa por fin a llamarse óvulo. El núcleo del óvulo maduro o pronúcleo femenino se ubica en el centro de la célula. La cabeza del espermatozoide se dilata formando el pronúcleo masculino. Ambos pronúcleos se encuentran en el centro del citoplasma, duplican su DNA y pierden su membrana nuclear. Los cromosomas del ahora llamado cigoto, que es diploide, se disponen en el huso mitótico para experimentar la primera mitosis. En resumen, las principales consecuencias de la fecundación son el restablecimiento de la dipliodía, determinación del sexo del embrión, dependiendo si es espermatozoide porta in cromosoma X o Y, y el inicio de la segmentación.
Segmentación
Luego de ocurrida la fecundación, el cigoto rodeado aún de la corona radiada y de la zona pelúcida, comienza a avanzar por la tuba uterina. Durante este trayecto se inicia la segmentación: una serie de divisiones sucesivas del cigoto sin un aumento total de tamaño, por lo que las células se van haciendo más pequeñas. Se identifica un cigoto de 2, 3, 4, 8 células, etc., también llamadas blastómeras. Cuando estas blastómeras se separan accidentalmente, generan gemelos monocigóticos. Una de las células del cigoto de dos blastómeras se divide antes y tenemos un cigoto de tres células por un período corto. Intresantemente esta célula contiene el sitio de entrada del espermatozoide y va a originar al embrión, mientras la célula que se divide retardadamente va a originar la placenta.
A partir de las 8 células el embrión en segmentación presenta el aspecto de una mora, por lo que se le denomina mórula. La mórula está rodeada por la zona pelúcida y se encuentra todavía en la tuba uterina. Las blastómeras de la mórula de 8 células comienzan un proceso de compactación por medio del establecimiento diferencial de uniones intercelulares entre sí. Se produce un estrecho sello entre las blastómeras, dejando en el interior de la mórula la masa celular interna formada por 3 células que presentan uniones comunicantes entre sí y que se encuentran aisladas del medio; quedando en el exterior la masa celular externa formada por 5 células que establecen uniones herméticas entre sí que se hallan en contacto con la tuba y sus secreciones. Las células de la masa celular interna proliferarán y formarán el embrioblasto, tejidos propios del embrión, y la masa celular externa proliferará dando origen al trofoblasto, que contribuirá a la formación de la placenta (Figura 2-4).
Cuando el embrión tiene 32 células se comienza a formar una cavidad en su interior, debido a la filtración de líquido desde la cavidad uterina, la que se denomina blastocele. En este momento, el embrión ha entrado en la fase de blastocisto. Las células de la masa celular interna se ubican hacia un polo, que se denominará el polo embrionario, mientras que las células de la masa celular externa se aplanan, forman gran parte de la pared del blastocisto y se disponen principalmente hacia el polo abembrionario. A fineales del cuarto día, cuando el blastocisto tiene alrededor de 100 células, se produce la eclosión, es decir el desprendimiento de la zona pelúcida y comienza la implantación.
Implantación
Aproximadamente en el quinto a sexto día post-fecundación comienza la implantación en la mucosa uterina. El útero tiene una forma triangular, alargada hacia el cuello uterino, una parte superior denominado fondo y dos paredes: una anterior y una posterior. La implantación ocurre normalmente en una de estas dos paredes, en la zona alta de la cavidad uterina. Si el blastocisto se implantara en el orificio interno del cérvix o en la tuba uterina, o en el peritoneo se estaría frente a implantaciones ectópicas, desarrollándose una placenta previa, embarazo tubario o embarazo abdominal respectivamente, que pueden generar complicaciones o aborto.
Cuando una mujer tiene una implantación en la pared anterior, normalmente en el segundo embarazo vuelve a implantarse su embrión en la misma zona, y si hay un tercer embarazo puede repetirse el fenómeno en la misma región.
La implantación busca que el embrión se anide en el endometrio. El embrión debe atravesar un epitelio endometrial indemne y debe superar la membrana basal de este epitelio para ingresar al tejido conectivo subyacente.
El proceso de implantación es simultáneo con el desarrollo del embrión bilaminar y ocurre durante la segunda semana de gestación. Al arribar el blastocisto a la cavidad uterina, alrededor del quinto día después de la fecundación, la zona pelúcida que ha mantenido la cohesión de las blastómeras y ha evitado la implantación ectópica en las trompas uterinas desaparece y las células adhesivas del trofoblasto comienzan a interactuar con el endometrio.
En el momento de la implantación el endometrio se encuentra en fase secretora o luteínica. Las glándulas están desarrolladas y las arteriolas de la submucosa se encuentran tortuosas y pletóricas En este período de máximo desarrollo de la mucosa es posible reconocer en ella tres capas:
1. la capa basal, delgada, que produce y renueva las capas que son más superficiales;
2. la capa esponjosa, intermedia, con abundantes conductos glandulares y vasos sanguíneos; y
3. una capa compacta, que es superficial y muy edematosa en esta fase del ciclo. La capa esponjosa y compacta constituyen la capa funcional de endometrio, que se renueva durante cada ciclo menstrual por la estimulación de los estrógenos secretados por las células foliculares y por la progesterona secretada por el cuerpo lúteo (Figura 2-5).
Durante la implantación el trofoblasto se diferencia en dos capas: el citotrofoblasto y el sincitiotrofoblasto. Hacia finales del sexto día de desarrollo el trofoblasto del polo embrionario se fija en la capa compacta del endometrio y comienza a profundizarse en la mucosa. El trofoblasto de esta región pierde sus límites y constituye un tejido multinucleado que no tiene límites claros llamado sincitiotrofoblasto. El sincitiotrofoblasto absorbe nutrientes, produce gonadotrofina coriónica y asume una acción histolítica, siendo el frente de invasión para permitir que el embrión que se introduzca en el estroma endometrial. El resto del trofoblasto, llamado citotrofoblasto, consiste en una capa de células ordenadas, con límites claros, que constituye principalmente la pared del blastocisto. Las mitosis del trofoblasto se producen principalmente en el citotrofoblasto por lo que las células del sincitiotrofoblasto tienen su origen en células del cito que van perdiendo sus membranas. A medida que la implantación progresa el sincitiotrofoblasto rodea gradualmente al blastocisto (Figuras 2-6a y b).
Al día nueve post fecundación el embrión se encuentra totalmente incluido en el endometrio y el epitelio endometrial presenta un coágulo de fibrina o tapón de cierre que sella el sitio de implantación. Paralelamente en la periferia del trofoblasto se observan una serie de capilares endometriales maternos congestionados de sangre.
Durante la implantación, el endometrio se autodestruye en respuesta a la interacción con el trofoblasto y experimenta la reacción decidual, donde se observa una hipertrofia celular endometrial y acumulación de glucógeno y lípidos en el citoplasma de las células endometriales. El endometrio que sufre reacción decidual pasa a llamarse decidua. La reacción decidual evita el rechazo del embrión, que es antigénicamente diferente a los tejidos maternos y, además, evita que la implantación progrese hasta el miometrio. Este proceso de anidamiento va progresando de manera que hacia el día 14 del desarrollo el blastocisto está sumergido en el endometrio y la capa compacta de la mucosa cubre el polo abembrionario del blastocisto.
A medida que el sincitiotrofoblasto prolifera, se forman espacios intercelulares, las lagunas trofoblásticas, que luego se conectarán con los capilares del endometrio, estableciéndose así la circulación placentaria, hecho que ocurre alrededor del día 12 del desarrollo. En estas lagunas se comienzan a proyectar las llamadas vellosidades coriónicas primarias formadas por sincitio y citotrofoblasto. Este hecho es capital debido a que el blastocisto ya ha alcanzado un tamaño cercano a 1 mm de diámetro, haciéndose difícil la nutrición del embrión sólo por el proceso de difusión de sustancias. Las blastómeras iniciales obtienen su nutrición desde el escaso vitelo presente en las reservas del óvulo. Luego la mórula utiliza los nutrientes presentes en las secreciones tubáricas y uterinas, desde el llamado embriotrofo o leche uterina. Más tarde, en la fase inicial de la implantación, el blastocisto utiliza los productos derivados de la destrucción del endometrio o histotrofo y, finalmente, la nutrición del embrión dependerá de la circulación placentaria o hemotrofo.
En el día 13, las lagunas trofoblásticas se encuentran llenas de sangre materna que proviene de la erosión de los capilares maternos del estroma endometrial, y estas lagunas constituyen el inicio de la circulación útero-placentaria. Estas lagunas se encuentran fuertemente congestionadas y es tal la presión, que puede haber algo de sangramiento a través de la región del coágulo de firina. Este sangramiento mediado por la implantación se confunde a veces con una menstruación.
Segunda semana de desarrollo - Disco bilaminar
El proceso de implantación se completa a mediados de la segunda semana, con un desarrollo notable del trofoblasto. Junto con estos cambios, el embrioblasto sufre modificaciones importantes que van a transformar este grupo de células en un disco celular bilaminar en la segunda semana y luego en disco trilaminar en la tercera semana.
Hacia el octavo día del desarrollo embrionario y en paralelo con el proceso de implantación, desde el embrioblasto se diferencian dos capas celulares: el epiblasto y el hipoblasto. El epiblasto, es una capa formada por células cilíndricas, altas, dispuestas radialmente que se diferencian de las células del embrioblasto que se encuentran revistiendo internamente al trofoblasto. El hipoblasto, es una capa formada por células cúbicas que están orientadas hacia la cavidad del blastocisto o blastocele. Por medio de la formación del epiblasto e hipoblasto la masa celular interna o embrioblasto se ha transformado en un disco bilaminar, con el epiblasto orientado al polo embrionario y el hipoblasto hacia el polo abembrionario (Figuras 2-6a y c).
En el día 8, junto con el proceso de implantación, entre las células del epiblasto se comienza a acumular líquido. Las células epiblásticas más cercanas al polo embrionario se aplanan y diferencian en amnioblastos, encargados de secretar el líquido amniótico. El conjunto de amnioblastos forma una membrana llamada amnios. Se origina así una nueva cavidad denominada cavidad amniótica entre el epiblasto y el amnios, que crece progresivamente.
Hacia el noveno día de desarrollo desde el hipoblasto surgen una membrana celular aplanada que se extiende sobre el citotrofoblasto cubriendo la cavidad del blastocisto. Esta capa célular recibe el nombre de endodermo extraemrionario, membrana exocelómica o membrana de Heuser, y la cavidad del blastocisto tapizada por esta membrana pasa a llamarse saco vitelino primario o cavidad exocelómica.
En este momento la cavidad amniótica contiene una pequeña cantidad de líquido y se encuentra entre el epiblasto y el citotrofoblasto. La cavidad exocelómica en tanto está limitada por la membrana de Heuser y por el hipoblasto. El disco embrionario bilaminar está ubicado entre el amnios y el saco vitelino primario (Figura 2-6b).
Hacia el décimo día, entre la membrana de Heuser y el citotrofoblasto se secreta una capa gruesa y reticulada y con muy pocas células de origen hipoblástico, que recibe el nombre de retículo extraembrionario. Hacia el día doce, en el retículo extraembrionario aparece una nueva población celular: el mesoderma extraembrionario. No es claro su mecanismo de formación pero se postula que desde el epiblasto del borde caudal del disco embrionario bilaminar se genera el mesoderma extraembrionario que se extiende en dos capas que bordean interna y externamente la capa reticular. Luego estas dos hojas crecen bordeando el amnios. Es decir, el mesodermo extraenbionario tiene el citotrofoblasto por fuera y amnios y la membrana exocelómica del por dentro. Luego, en el interior del reticulado se crean espacios que van progresivamente convergiendo de modo tal que en el mesoderma extraembrionario se reconocen dos hojas: una hoja unida al citotrofoblasto y al amnios, la somatopleura extraembrionaria u hoja parietal del mesoderma extraembrionario y otra hoja adherida al saco vitelino: la esplacnopleura extraembrionaria u hoja visceral del mesoderma extraembrionario. Hacia el décimo tercer día de desarrollo el espacio entre la somatopleura y la esplacnopleura ha crecido y éste es el que recibe el nombre de cavidad coriónica o celoma extraembrionario (Figuras 2-7a y b).
En particular, la somatopleura que cubre la superficie del trofoblasto o sea la que se encuentra en la pared de la cavidad coriónica recibe el nombre de lámina coriónica y estas dos hojas, la somatopleura más el trofoblasto forman el anexo embrionario llamado corion. Así, la cavidad amniótica se encuentra delimitada por el amnios y la cavidad coriónica se encuentra delimitada por el corion.
La cavidad coriónica sigue expandiéndose de modo que al día 15 el embrión con su cavidad amniótica y su saco vitelino adyacentes queda suspendido por un tallo de somatopleura extraembrionaria llamado pedículo de fijación. A esta edad ya se ha definido una polaridad en el embrión, de modo que el pedículo de fijación une el extremo caudal del disco bilaminar y el corion. El pedículo de fijación formará el cordón umbilical en el período fetal.
Concomitantemente con estos procesos, al día 12 una nueva oleada de células migran desde el hipoblasto hacia el interior del saco vitelino primario para formar una nueva cavidad más pequeña: el saco vitelino secundario. Las células de origen hipoblástico que lo recubren internamente conforman el endodermo extraembrionario. La membrana exocelómica y la cavidad exocelómica son empujadas en este proceso hacia el fondo de la cavidad coriónica, formando los llamados quistes exocelómicos, que al decimotercer día se pueden observar en el fondo de la cavidad coriónica, donde terminan por degenerar.
Si bien es cierto que el saco vitelino es un elemento vestigial en el hombre no es menos importante; ya que a través de él migran, como vimos en el capítulo anterior, las células germinales primordiales y en el mesoderma extraembrionario de su pared se diferencian los primeros vasos sanguíneos hacia el día 17-18.
Tercera semana de desarrollo - Disco trilaminar
Durante la tercera semana de vida, en el embrión bilaminar, constituido por epiblasto e hipoblasto, ocurre el proceso de gastrulación, definido como una serie de movimientos celulares que darán origen las tres capas germinativas del embrión: ectodermo, mesodermo y endodermo, generando así un disco embrionario trilaminar.
Hacia el día 15 de desarrollo se observan varios elementos que nos permiten definir los ejes del embrión: aparece un surco en la línea media caudal del epiblasto, que es este momento aparece elongado y ensanchado en su porción cefálica: es el llamado surco primitivo. Al día 16, este surco primitivo llega a ocupar la mitad del disco embrionario y en su extremo cefálico de este surco se observa una depresión, la llamada fosita primitiva que se continúa hacia cefálico con un reborde denominado nodo primitivo o de Hensen. En su conjunto, nodo, fosita y surco primitivo reciben el nombre de línea primitiva. A estas alturas el embrión tiene forma de una paleta con el extremo cefálico ancho y la región caudal angosta. En el extremo cefálico del embrión en tanto, las células hipoblásticas en un área circular limitada adoptan una disposición columnar, estableciendo una estrecha unión con el epiblasto suprayacente. Esta zona denominada membrana bucofaríngea marca el sitio de la futura cavidad bucal. En el extremo caudal de la línea primitiva se hace evidente un área circular, la membrana cloacal, similar pero más reducida que la membrana bucofarínge y marca el sitio de la futura región anal. Cuando esta membrana se forma, lo que ocurre hacia el día 16, el saco vitelino emite una pequeña extensión hacia el pedículo de fijación que recibe el nombre de alantoides. En la membrana bucofaríngea y cloacal existe una firme unión entre ectodermo y endodermo.
Ocurre así la gastrulación, la que se puede dividir didácticamente en tres movimientos que se sobreponen. El primer movimiento de la gastrulación formará el endodermo, el segundo movimiento de la gastrulación formará el mesodermo y el tercer movimiento formará la placa precordal y la notocorda (Figuras 2-8a y b).
Primer movimiento de la gastrulación
Hacia el día 16 células del epiblasto proliferan, migran hacia la línea media y se invaginan por el surco primitivo para luego extenderse hacia lateral desplazando al hipoblasto, originando el endodermo embrinario.
Segundo movimiento de la gastrulación
Una segunda oleada de células epiblásticas se invaginan por el surco primitivo para intercalarse entre el epiblasto y el endodermo reción formado, originando así el mesodermo. Las células que quedan en el epiblasto pasarán en este momento a llamarse ectodermo (Figura 2-9).
Tercer movimiento de la gastrulación
Células en la región del nodo primitivo comienzan a invaginarse por la fosita primitiva y migran a nivel de la línea media formando la placa precordal por cefálico y el proceso notocordal por caudal. La placa precordal, masa compacta de mesodermo, se sitúa por detrás de la membrana bucofaríngea y constituye en un importante centro organizador de la región cefálica del embrión. El proceso notocordal tiene su origen en las células epiblásticas prenotocordales. Este proceso es primero un tubo hueco, luego se abre y fusiona con el endodermo para formar la placa notocordal y finalmente se levanta transformado en una cuerda sólida: la notocorda (Figura 2-8b).
La notocorda ejerce un efecto inductor morfogenético, en base a interacciones moleculares, sobre el ectodermo suprayacente el cual aumenta de grosor formando la llamada placa neural que se extiende desde el nodo primitivo hasta la lámina precordal. La notocorda servirá además de núcleo organizador del esqueleto axial.
Derivados de las hojas embrionarias
En esta etapa del desarrollo, la totipotencialidad presente en las blastómeras iniciales se ha reducido notablemente y cada una de estas hojas embrionarias dará origen a diferentes tejidos en el embrión. Así, por ejemplo, el ectodermo formado por células epiteliales columnares da origen a:
1. sistema nervioso central y periférico,
2. epidermis, pelos y uñas, y
3. esmalte dentario.
El mesodermo formado por células reticulares con abundante matriz extracelular da origen a:
1. dermis, cartílago y hueso,
2. musculatura lisa y estriada,
3. corazón, bazo, vasos sanguíneos y linfáticos,
4. células sanguíneas, y
5. gónadas y riñón.
El endodermo formado por celulas epiteliales planas da origen a:
1. epitelio del tracto digestivo y respiratorio,
2. epitelio de vejiga y uretra,
3. tiroides, paratiroides, hígado y páncreas, y
4. amígdalas y timo.
Mediante investigación de trazado de linaje se ha podido establecer un mapa de los territorios presuntivos del epiblasto que darán origen a diferentes linajes celulares. Los factores determinantes de estos linajes celulares parecen estar condicionados por una clave temporal, es decir el momento en que las células epiblásticas migran hacia la línea primitiva, y por una clave topológica, es decir el lugar por donde migran dichas células para establecerse en el disco trilaminar.
Segmentación del mesodermo
Hacia el día 16, el recientemente creado mesodermo intraembrionario adopta la disposición de una lámina delgada. Hacia el día 17 el mesodermo se segmenta en porciones: 1) Mesodermo axial, constituido por la notocorda y la placa precordal 2) El mesodermo paraxial, próximo a la notocorda. 3) El mesodermo intermedio, por fuera del anterior y 4) El mesodermo lateral, hacia el borde del disco embrionario. Hacia fines de la tercera semana el mesodermo paraxial de la región cervical prolifera y forma relieves simétricos visibles a los lados del tubo neural. Estos relieves reciben el nombre de somitos. La formación de los somitos continúa progresivamente hacia caudal. En tanto, hacia la mitad de la tercera semana del desarrollo, células mesodérmicas ubicadas por delante y hacia lateral de la lámina precordal conforman el área cardiogénica en la cual se diferencian formando los primeros vasos sanguíneos intraembrionarios que rápidamente se extienden por el embrión. En el área cardiogénica se forma un tubo vascular doble, el tubo cardíaco, que dará origen al corazón (Figuras 2-10a, b y 2-11).
Vellosidades coriónicas
A finales de la segunda semana, en el corion comienzan a desarrollarse las vellosidades coriónicas. Se desarrollan primero las vellosidades coriónicas primarias, formadas por citotrofoblasto más sincitiotrofoblasto. A comienzos de la tercera semana, la lámina coriónica del mesoderma extraembrionario o somatopleura extraembrionaria prolifera en el interior de las vellosidades coriónicas primarias, formando un centro de tejido laxo, generándo así las vellosidades coriónicas secundarias. Hacia finales de la tercera semana algunas células mesodérmicas darán origen a vasos sanguíneos en el interior de las vellosidades conformando las vellosidades coriónicas terciarias. Pronto estos vasos sanguíneos se conectarán con la circulación del embrión por medio de los vasos umbilicales ubicados en el pedículo de fijación, estableciéndose la circulación materno-fetal. De esta manera que el sistema cardiovascular es el primer sistema funcionante del organismo (Figura 2-12).
Período embrionario - Tercera a octava semana
Durante estas cinco semanas se produce el desarrollo de todos los sistemas corporales, aun cuando algunos sistemas como el nervioso y el cardiovascular ya estaban esbozados a mediados de la tercera semana. Hacia el final de este período el embrión presenta un aspecto más humano. Este proceso de morfogénesis involucra una serie de interacciones tisulares complejas en una secuencia temporal precisa, razón por la cual en este período la exposición de embriones a teratógenos (agentes que producen alteraciones del desarrollo) como radiaciones (rayos x), virus (rubéola) o fármacos (talidomida) provoca malformaciones congénitas mayores.
Desarrollo del mesodermo
Como se ha señalado anteriormente, el recién formado mesoderma intraembrionario se ha segmentado en: mesoderma paraaxial, colocado al lado de la notocorda; mesoderma intermedio, ubicado más hacia lateral y; el mesoderma lateral, en relación con el borde del disco embrionario (Figura 2-10b).
Desarrollo del mesodermo paraaxial
Hacia el final de la tercera semana de desarrollo se hacen evidente en el aspecto dorsal del embrión, a ambos lados del tubo neural, la presencia de los somitos. Estos somitos, que corresponden a engrosamientos del mesoderma paraaxial, presentan una secuencia de aparición precisa, de manera que es posible determinar la edad del embrión conociendo el número de somitos. Los primeros pares de somitos aparecen el día 20 en la región cervical del embrión y continúan apareciendo de a tres pares por día hasta alcanzar un número de 42 a 44 pares.
Así, es posible determinar la edad del embrión de acuerdo a la Tabla 2-1.
Tabla 2-1
Determinar edad del embrión
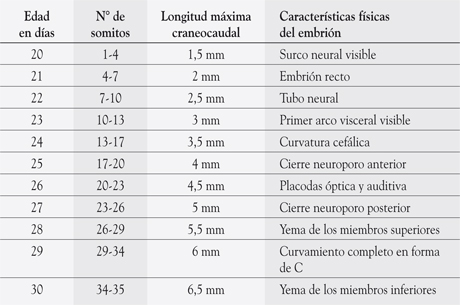
Los pares de somitos se denominan según su ubicación: 4 occipitales (de los cuales el primer par involuciona), 8 cervicales, 12 torácicos, 5 lumbares, 5 sacros y 8 a 10 coxígeos (de los cuales involucionan los más caudales).
Hacia el comienzo de la cuarta semana las células mesodérmicas de los somitos van a diferenciarse en tres variedades. Las células de la zona medial del somito rodean al tubo neural y la notocorda formando el esclerotoma, tejido que dará origen a los elementos óseos del esqueleto axial (vértebras y costillas) del sector correspondiente del somito. Las células de la región dorsal de los somitos van a diferenciar el miotoma, tejido que dará origen a la musculatura estriada del sector correspondiente; y el dermatoma, tejido que da origen a la dermis y tejido subcutáneo del área correspondiente. Cada somito recibe una rama arterial y una rama nerviosa que se distribuyen precisamente por los tejidos que dan origen (Figura 2-13).
En relación a las vértebras, del esclerotoma va a condensar primero el tejido alrededor de la notocorda para formar el cuerpo vertebral y luego se condensa hacia posterior para rodear al tubo neural, formando el canal vertebral que protegerá a la futura médula espinal.
Cuando hacen su aparición los somitos caudales, los somitos ubicados más hacia cefálico ya se han diferenciado de modo tal que en ningún momento es posible observar los aproximadamente 42 pares de somitos simultáneamente. Esta forma de ordenamiento del cuerpo del embrión en unidades homólogas, llamadas metámeras, dispuestas en el eje céfalo-caudal, se hará especialmente notorio después del nacimiento en el tronco donde se aprecia la disposición secuencial de las vértebras, costillas y arterias, nervios y músculos intercostales.
Desarrollo del mesodermo intermedio
Desde el mesodermo intermedio de los dos lados del cuerpo surgen estructuras renales secuenciales y de organización más compleja. En él se distinguen los nefrotomos, cúmulos de células que se segmentan a nivel cervical y torácico y el cordón nefrógeno, masa no segmentada que formará las unidades excretoras definitivas del sistema urinario y participará en la formación de las gónadas.
Al día 24, de la región cervical y torácica alta se forma el pronefros que consiste en acúmulos llamados nefrotomos segmentarios, transitorios, no funcionales y que rápidamente involucionan. Cuando el pronefros regresa durante la cuarta semana surge el mesonefros: masa no segmentada de mesodermo intermedio de la región torácica baja y lumbar alta. El mesonefros diferencia nefrones completos y simples que funcionan transitoriamente y luego degenera. Sin embargo, el conducto mesonéfrico persiste en el embrión masculino y formará el ducto deferente. Finalmente, el mesodermo intermedio de la región lumbar baja y sacra diferencia el metanefros, desde donde surgirán las unidades excretoras del riñón definitivo, alrededor de la quinta semana de vida embrionaria.
Desarrollo del mesodermo lateral
El mesodermo lateral ubicado hacia el borde del disco embrionario se divide en dos hojas una que se extiende hacia el amnios, la llamada somatopleura intraembrionaria u hoja parietal del mesodermo intraembrionario, y otra que se extiende hacia el saco vitelino secundario, la esplacnopleura intraembrionaria u hoja visceral del mesodermo intraembrionario. A cada lado en los bordes del disco embrionario, entre estas dos hojas somática y esplácnica del mesodermo, se irá formando progresivamente un espacio que recibe el nombre de celoma intraembrionario, el cual comunica hacia lateral con el celoma extraembrionario. La somatoplaura intraembrionaria dará origen a las membranas serosas: pleura, pericardio y peritoneo, que envuelven al pulmón, corazón y abdomen respectivamente. La esplacnopleura intraembrionaria formará la pared muscular del tubo digestivo y de la vía respiratoria baja. Posteriormente el celoma intraembrionario dará origen a las cavidades corporales pericárdica, pleural y peritoneal (Figura 2-10b).
Neurulación
El ectodermo recién formado durante la gastulación recibe inducciones de la notocorda subyacente para diferencial en el ectodermo un neuroepitelio, también llamado neuroectodermo, que comienza a engrosarse formando la placa neural. Hacia el día 19 comienza la neurulación, que es el proceso por el cual el neuroepitelio de la placa neural se convierte en tubo neural. Durante el día 20 los bordes de la placa neural se levantan formando los pliegues neurales que determinan la aparición del surco neural en la línea media. El surco neural se oserva inmediatamente por delante de la línea primitiva. Los pliegues neurales son especialmente prominentes en el extremo craneal del embrión pues en esa región se desarrollará el encéfalo. Hacia el final de la tercera semana los pliegues neurales se aproximan y se fusionan transformando la placa neural en un tubo neural que contiene el canal neural. El proceso de cierre comienza al día 22 del desarrollo, a la altura de la futura región cervical del feto. A medida que el tubo se cierra se va profundizando el ectodermo lo cubre y se diferencia en la epidermis de la región dorsal. Esta epidermis es continua con el resto de la epidermis diferenciada del ectodermo y va rodeando a todo el embrión gracias a un proceso de plegamiento embrionario. El borde del pliegue neural presenta una estructura especializada llamada cresta neural. De las células de la cresta neural se originarán todas las neuronas y glías del sistema nervioso periférico, las células de la médula suprarrenal y los melanocitos; en tanto desde el tubo neural se originan neuronas y glías del sistema nervioso central, lo que incluye cerebro, tronco encefálico, cerebelo y médula espinal (Figura 2-14).
En el inicio de la cuarta semana ha comenzado el cierre del tubo neural, proceso que se inicia a la altura del cuarto somito cervical y se extiende rápidamente hacia cefálico y hacia caudal. Los extremos del tubo neural, llamados neuroporo anterior el cefálico y neuroporo posterior el caudal, permanecen temporalmente abiertos y comunicados con la cavidad amniótica. Hacia el día 25 (período de 18 a 20 somitos) se produce el cierre del neuroporo anterior y el día 27 (período de 25 somitos) lo hace el neuroporo posterior (Figura 2-13).
El cierre de los neuroporos son fundamentales para el correcto desarrollo del sistema nervioso; la falta de cierre del neuroporo craneal origina anencefalia, ausencia de desarrollo del encéfalo. Mientras que la falta de cierre del neuroporo caudal origina una anomalía llamada espina bífida. Hacia el día 22-23 el extremo cefálico del embrión comienza a desarrollar una importante flexión ventral. En esta región se desarrollará el cerebro medio o mesencéfalo. Por delante se desarrollará el cerebro anterior o procencéfalo y por detrás el cerebro posterior o rombencéfalo.
De esta manera se completa el proceso de neurulación y el sistema nervioso central está formado por una porción caudal, tubular y estrecha, la futura médula espinal; y una porción cefálica, dilatada en varias regiones llamadas las vesículas cerebrales, que incluyen las porciones ya mencionadas: prosencéfalo, mesencéfalo y rombencéfalo.
En el momento del cierre del tubo neural, aproximadamente al día 26, en la región cefálica del embrión se hacen evidentes dos engrosamientos ectodérmicos: la placoda del cristalino y la placoda auditiva; las que invaginarán diferenciando posteriormente la vesícula óptica y la vesícula auditiva respectivamente (Figura 2-15).
Desarrollo del plegamiento del embrión
El desarrollo excepcional que presentan el tubo neural y las vesículas cerebrales afecta a la forma del embrión, determinando la aparición de curvaturas en el eje longitudinal y transversal de él. En el eje longitudinal, se hacen evidentes: 1) una curvatura cefálica, donde el encéfalo en desarrollo se proyecta por delante de la lámina precordal y del área cardiogénica, desplazando a éstas hacia abajo y hacia la superficie ventral del embrión. En éste movimiento, parte del saco vitelino queda incorporado en el interior del embrión formando el intestino anterior. 2) una curvatura caudal, producida por el desarrollo de la médula espinal, la cual se extiende más allá de la membrana cloacal, desplazando a ésta última hacia una posición más ventral. Durante este movimiento parte del saco vitelino es incorporado al interior del embrión formando el intestino posterior. En el eje transversal, causado principalmente por el desarrollo de los somitos, se produce el plegamiento lateral del embrión, de modo tal que los bordes del embrión se proyectan hacia ventral y la hoja somática o parietal del mesodermo se va cerrando en la línea media, conformando así las paredes corporales anterolaterales. Este movimiento reduce cada vez más al saco vitelino y la zona media de él conformará el intestino medio. El intestino medio guarda una estrecha comunicación con el saco vitelino que recibe el nombre de conducto onfalomesentérico. En resumen, el endodermo junto una porción del saco vitelino forma el tubo intestinal que recorre todo el embrión desde la membrana bucofaríngea a la membrana cloacal. El intestino anterior formará esófago, estómago y duodeno proximal, además originará también un divertículo que se extenderá y ramificará para formar el epitelio respiratorio. Del duodeno proximal se formará el hígado, vesícula biliar y páncreas. El intestino medio produce el duodeno distal, el yeyuno, íleon, ciego, colon ascendente y los dos tercios proximales del colon transverso. Del intestino posterior derivan el tercio distal del colon transverso, el colon descendente, el colon sigmoideo y el recto.
En la medida que el saco vitelino se reduce, el amnios crece y literalmente va englobando al embrión, el cual queda flotando en el líquido amniótico. Del mismo modo, el pedículo de fijación que inicialmente se ubicaba en el extremo caudal, se va progresivamente desplazando hacia la superficie ventral del embrión, y su punto de inserción se reduce a una zona angosta en la región umbilical. El amnios formará la cubierta externa del cordón umbilical, dentro del cual quedan incluidos el saco vitelino y el alantoides. El crecimiento del amnios reduce progresivamente el celoma extraembrionario o cavidad coriónica, la que desaparece hacia el final de segundo mes de gestación (Figuras 2-16a, b y c).
Hernia umbilical fisiológica
Como consecuencia del rápido crecimiento del intestino y del desarrollo del hígado, la cavidad abdominal se hace estrecha para contener a las asas intestinales las que se ubican temporalmente en el interior del cordón umbilical, desarrollándose en el embrión de seis semanas la hernia umbilical fisiológica. En el punto de unión del cordón umbilical a la pared abdominal del feto existe una amplia comunicación entre el celoma intraembrionario y el extraembrionario, donde se alojan transitoriamente las vísceras digestivas. A medida que se desarrolla el embrión esta comunicación se reducirá progresivamente, por el crecimiento de la pared corporal (Figura 2-16b).
Desarrollo de los arcos viscerales
Hacia el final de la cuarta semana y concomitantemente con el desarrollo de las vesículas cerebrales y la formación de la curvatura cefálica aparecen, en la región cefálica del embrión, cuatro pares de rodetes transversales, los arcos branquiales también llamados faríngeos o viscerales, que se ubican hacia caudal del estomodeo o primitiva cavidad bucal. Estos arcos faríngeos, formados por tejido mesenquimático originado desde las crestas neurales de las vesículas cerebrales, están separados en la superficie externa del embrión por las hendiduras faríngeas y en el aspecto interno por las bolsas faríngeas. Los arcos faríngeos dan origen a huesos y músculos del territorio maxilofacial y cervical además de participar en la formación de la lengua, faringe y laringe. La primera hendidura faríngea participa en la formación del conducto auditivo externo y del tímpano. Las bolsas faríngeas participan en la formación de oído medio, glándula tiroides, paratiroides, timo, cavidad timpánica, conducto faringo-timpánico y amígdala palatina. A medida que los arcos faríngeos se desarrollan se va esbozando la cara y cuello del embrión (Figura 2-17).
Desarrollo de los miembros superiores e inferiores
En paralelo con los cambios registrados en la región cefálica, al comienzo de la quinta semana aparecen, en el aspecto lateral del embrión, los esbozos de los miembros superiores e inferiores, como yemas semejantes a palas de remo. Los miembros superiores se ubican detrás de prominencia cardíaca, entre el cuarto somito cervical y el primer somito torácico (cuyos nervios, formando el plexo braquial, la inervarán); y en su desarrollo van adelantadas respecto de los miembros inferiores. Estas últimas se ubican caudal al punto de inserción del pedículo umbilical entre los somitos lumbares y los primeros somitos sacros (de donde proviene su inervación por medio del plexo lumbosacro). En el desarrollo de los miembros superiores e inferiores se forman primero los segmentos más distales (mano y pie) y posteriormente los segmentos más proximales, separados por constricciones circulares que los demarcan. En la futura mano y pie, aparecen surcos radiales que progresivamente irán delimitando a los dedos del miembro superior. Inicialmente los miembros superiores e inferiores sobresalen perpendicularmente a los lados del cuerpo pero después sufren un cambio de orientación, de modo que el miembro superior rota 45° hacia dorsal y la inferior lo hace 45° hacia ventral, de manera que se establece una diferencia de 90°, quedando el codo orientado hacia atrás y la rodilla hacia adelante.
Inicialmente, los dedos se encuentran unidos por la membrana interdigital como las extremidades de una rana, elemento que se adelgaza y desaparece, permitiendo identificar cada dedo. La falla en la separación de los dedos produce sindactilia o dedos fusionados (Figura 2-18).
Características del embrión entre la cuarta y octava semana
• Cuarta semana. Al inicio de la cuarta semana el embrión es recto y en el aspecto dorsal se aprecia el relieve de los somitos. Hacia la mitad de la cuarta semana, en el aspecto ventral aparecen los arcos branquiales y comienza a incurvarse el cuerpo del embrión. Al final de la cuarta semana, en el aspecto lateral se aprecian las yemas de los miembros superiores e inferiores y en la región cefálica se observan las placodas ópticas y auditivas.
• Quinta semana. Se observa un desarrollo notable del encéfalo. La cara del embrión toma contacto con la prominencia cardíaca. El segundo arco visceral crece formando el opérculo, y cubre a los arcos más caudales, formándose una depresión ectodérmica llamada seno cervical. Ecográficamente a esta edad es posible visualizar el saco gestacional, como una imagen quística más o menos esférica.
• Sexta semana. En los miembros superiores se identifican las regiones del codo y la muñeca y las placas de las manos. En la cabeza, en relación con la primera hendidura faríngea, comienza a formarse el pabellón auricular. Ecográficamente a esta edad es posible visualizar en el interior del saco gestacional al embrión, como una pequeña zona ecorefringente.
• Séptima semana. El intestino en desarrollo se proyecta en el cordón umbilical, formando la hernia umbilical fisiológica. En las manos se observa los rayos digitales que van delimitando a los dedos. Ecográficamente a esta edad el saco gestacional aparece de mayor tamaño y en su interior se aprecia claramente al embrión; también es posible advertir, en la ecografías de tiempo real, sus movimientos cardíacos.
• Octava semana. Se distinguen los dedos de manos y pies y la cola ha involucionado. El embrión ha adquirido características morfológicas humanas. La cabeza representa la mitad de la longitud del embrión y los párpados se desarrollan y se fusionan hacia el final de ésta semana. Ecográficamente a esta edad se aprecia en el interior del saco gestacional el embrión y el saco vitelino. Además se observan los esbozos de los miembros superiores e inferiores y se pueden visualizar los movimientos bruscos del embrión.
Estimación de la edad gestacional
Existen dos métodos diferentes para determinar la fecha del embarazo. La edad concepcional o de fecundación, utilizada por los embriólogos, fecha el embarazo a partir de la fecundación; dato que es exacto sólo en pacientes que se han sometido a fertilización o inseminación artificial. La edad menstrual, utilizada por los obstetras, fecha el embarazo desde el primer día del último período menstrual (FUR = fecha de la última regla) de la paciente. La edad menstrual del embrión es dos semanas más que la edad concepcional y, en consecuencia, se debe restar ± 14 días a la FUR para obtener la edad de fecundación. Así, a un embrión de edad menstrual de ocho semanas le corresponde una edad concepcional de seis semanas, y la duración del embarazo es de 40 semanas de edad menstrual y de 38 semanas de edad concepcional. La estimación de la edad gestacional a partir de la historia menstrual de la mujer no está exenta de error. Por ejemplo, la probabilidad de error en pacientes que se embarazan después de suprimir los anticonceptivos orales es alta, ya que es muy variable el intervalo entre la suspensión de las hormonas y el inicio de la ovulación. Otra condición que puede inducir a error es el discreto sangrado que se presenta en algunos casos durante la implantación del embrión, lo que es interpretado como flujo menstrual.
Para determinar la edad del embrión se utiliza el recuento de los somitos y las características del desarrollo de los miembros superiores e inferiores. También es posible determinar la edad embrionaria a partir de la longitud cráneo-glútea (o coronilla rabadilla). Así, un embrión de 5 semanas mide entre 5 a 8 mm, de 6 semanas mide 10 a 14 mm, de 7 semanas mide 17 a 22 mm y de 8 semanas mide 28 a 30 mm.
Período fetal y placenta - Tercer mes al nacimiento
El período fetal se caracteriza por el crecimiento y maduración de los tejidos y órganos que se han diferenciado durante el período de organogénesis o embrionario, analizado en el capítulo precedente. Durante el período fetal la cabeza crece más lentamente que el cuerpo de modo que se modifica la relación cráneo/cuerpo, adquiriendo el feto proporciones más armónicas; observándose un incremento sostenido de la talla y del peso.
Períodos del embarazo
Los embriólogos dividen la gestación en períodos desiguales que toman como base hitos relevantes del desarrollo. Así, se describen:
• Período de desarrollo precoz, que va desde la fecundación hasta la tercera semana, donde ocurre la segmentación, implantación y gastrulación. En este lapso, la exposición a agentes teratógenos induce el aborto.
• Período embrionario, que se extiende entre la cuarta y octava semanas, donde ocurre la organogénesis de los sistemas corporales. Etapa de máxima vulnerabilidad a agentes teratógenos, generándose malformaciones mayores.
• Período fetal, que va desde el tercer al noveno mes, donde se verifica el crecimiento y maduración funcional de los órganos y sistemas. Lapso de menor vulnerabilidad a agentes teratógenos.
Los obstetras en cambio dividen al embarazo en tres períodos iguales:
• Primer trimestre: al final del cual ya están desarrollados todos los sistemas mayores del organismo. Período de mayor susceptibilidad a los agentes que provocan anomalías del desarrollo.
• Segundo trimestre: período durante el cual el feto adquiere un tamaño suficiente, de modo que es posible observar mediante ecografía buenos detalles anatómicos y detectar posibles anomalías. Hacia fines del segundo trimestre el feto puede sobrevivir si nace prematuramente.
• Tercer trimestre: período durante el cual el feto presenta una ganancia de talla y de peso considerable. En esta etapa el feto suele sobrevivir si nace prematuramente.
Desarrollo fetal
Tercer mes
Durante el tercer mes, el rostro adquiere un aspecto más humano debido a que los ojos adoptan una posición más frontal y los pabellones auriculares, inicialmente colocados en la zona cervical, se ubican en la región lateral de la cabeza. A esta edad gestacional, los párpados están fusionados, aparecen los centros de osificación en los huesos largos y en la base del cráneo. Hacia fines del tercer mes los genitales han alcanzado un desarrollo suficiente como para poder definir el sexo del feto mediante ecografía. En este período es posible determinar ecográficamente el diámetro biparietal, dato que servirá de referencia para hacer el seguimiento del desarrollo fetal. La pared anterolateral del abdomen ha crecido lo suficiente como para incorporar en la cavidad abdominal las asas intestinales, reduciéndose así la hernia umbilical fisiológica (Figura 2-19).
Cuarto mes
Durante el cuarto mes el feto crece rápidamente, aun cuando el peso se incrementa lentamente, de manera tal que a mediados del período gestacional tiene la mitad de la talla de un recién nacido, pero pesa menos de 500 gramos. Al cuarto mes la piel del feto está cubierta por un vello fino, el lanugo, y son visibles las cejas y el cabello. Ecográficamente en esta etapa es posible identificar el cordón umbilical y visualizar en él la vena y las arterias umbilicales, siguiendo su trayecto en el abdomen del feto (Figuras 2-20a y b).
Quinto mes
En el quinto mes, los miembros inferiores crecen y adquieren una proporción más armónica. A esta edad la madre percibe los movimientos fetales y es posible auscultar los latidos fetales. La piel está cubierta por una secreción grasosa llamada unto sebáceo o vérnix caseosa. Esta capa grasosa evita la maceración de la piel del feto que flota en el líquido amniótico. En este período se canalizan los canales anal y vaginal, y comienza la migración del testículo. Ecográficamente ya son visibles los riñones y la vejiga urinaria (Figuras 2-21a y b).
Sexto mes
Durante el sexto mes se produce la madurez pulmonar y las células alveolares producen surfactante, factor tenso activo que permite la permeabilidad alveolar. La piel es delgada y arrugada por la falta de grasa subcutánea, y se aprecian las uñas de los dedos de las manos. En este período es posible generar algunos reflejos fetales (movimientos bruscos) al aplicar ruidos vibroacústico en el abdomen de la madre. Ecográficamente, el perímetro craneal de circular que se observaba en los meses precedentes, se aprecia ahora de forma oval (Figura 2-22).
Séptimo mes
Durante el séptimo mes, el sistema nervioso central ha madurado de manera que puede controlar los movimientos respiratorios y la temperatura corporal. Los pulmones son capaces de respirar, aparecen las uñas en los dedos de los pies y comienza el depósito de grasa subcutánea, lo que borra las arrugas de la piel. Ecográficamente, es posible identificar el diafragma, cuyo espesor es de 2 a 3 mm (Figura 2-23).
Octavo mes
Durante el octavo mes, hay un incremento de la grasa subcutánea, lo que hace que aparezcan redondeados los contornos del feto, dándoles un aspecto rechoncho. Esto es posible de observar en las ecografías, especialmente en el abdomen fetal, donde el panículo adiposo tiene alrededor de 1 cm de espesor. Los testículos ya han cruzado el canal inguinal y se aproximan a la bolsa escrotal. El perímetro craneal y el abdominal son más o menos similares (Figuras 2-22 y 2-24).
Noveno mes
Durante el noveno mes, el feto presenta reflejo de prehensión y de orientación a la luz. Se mantiene el depósito de grasa subcutánea y el perímetro craneal representa la región de mayor circunferencia corporal. El feto de término pesa alrededor de 3 kilos, con una talla de 50 cm, medida desde el cráneo al talón. Generalmente, los fetos masculinos miden y pesan más que los fetos femeninos. Ecográficamente, es posible observar núcleos de osificación en el miembro inferior, especialmente en el extremo distal del fémur y en el extremo proximal de la tibia (Figuras 2-25a, b y c).
Determinación de la talla fetal
Para determinar la talla fetal se emplean las siguientes fórmulas:
• Para los cuatro primeros meses de gestación: longitud = edad al cuadrado, donde la longitud es expresada en cm y medida entre cráneo y talón; y la edad es expresada en meses. Por ejemplo, un feto de 3 meses mide 9 cm.
• Para la segunda mitad del embarazo: longitud = edad x 5 + 5, donde la longitud es expresada en cm y medida entre cráneo y talón; y la edad es expresada en meses. Así, un feto de 7 meses mide 40 cm. Durante éste período, el feto es menos vulnerable debido a que todos los sistemas corporales ya estaban diferenciados en el período prefetal. Sin embargo, el crecimiento fetal puede ser seriamente afectado por factores maternos. Así, el tabaquismo produce fetos pequeños; la desnutrición materna produce fetos de bajo peso, y la combinación de ambos tiene un efecto sumativo. El uso de marihuana durante el embarazo provoca retardo del crecimiento y retardo mental leve en el feto. El alcoholismo materno tiene efectos devastadores sobre el feto, el cual presenta retardo del crecimiento pre y postnatal, retardo mental y anomalías congénitas como hipoplasia maxilar y cardiopatías congénitas, estas alteraciones son más marcadas si se acompañan, como generalmente ocurre, con malnutrición materna. Es importante consignar que el consumo de 70 ml (medio vaso) de licor diario produce efectos alcohólicos en el feto. Otros factores que afectan el crecimiento fetal son: El embarazo múltiple, donde dos o más fetos compiten por la nutrición disponible a través de la placenta. Alteraciones del flujo sanguíneo placentario, como infartos o desprendimientos de la placenta. Factores genéticos como ocurre en fetos portadores de trisomía del cromosoma 21 (síndrome de Down) y trisomía del cromosoma 18.
Membranas fetales
Al inicio del segundo mes, el trofoblasto presenta abundantes vellosidades coriónicas secundarias y terciarias que le dan un aspecto radiado. Las vellosidades se extienden entre la placa coriónica y la envoltura citotrofoblástica. Esta envoltura o concha citotrofoblástica corresponde a tejido trofoblástico que separa los tejidos fetales de la decidua, nombre que recibe la capa funcional del endometrio gravídico, la que será expulsada durante el alumbramiento. Como vimos en la unidad 2, las vellosidades primarias están formadas por sincitio y citotrofoblasto. Las vellosidades secundarias presentan además un centro de mesoderma extraembrionario y las vellosidades terciarias contienen vasos sanguíneos en su interior. De acuerdo a su extensión, se distinguen vellosidades libres, las cuales no se fijan en la concha citotrofoblástica y flotan en el espacio intervelloso o cámara hemática, bañadas por la sangre materna. En cambio, las vellosidades ancla llegan hasta la envoltura citotrofoblástica y a través de ella hasta la decidua, permitiendo la fijación del embrión.
En las primeras semanas del desarrollo, las vellosidades cubren toda la superficie del corion. Posteriormente, las vellosidades del polo embrionario se desarrollan dando origen al corion frondoso, mientras que las vellosidades del polo abembrionario se reducen y hacia el tercer mes, esta parte recibe el nombre de corion liso (Figuras 2-12 y 2-26).
Endometrio gravídico o decidua
La decidua que se relaciona con el corion frondoso se denomina decidua basal; la decidua que cubre el polo abembrionario se llama decidua capsular; la decidua que cubre las paredes del útero se denomina decidua parietal. Con el crecimiento del feto, la decidua capsular y parietal se fusionan desapareciendo la cavidad uterina. A medida que crece el amnios, se reduce concomitantemente la cavidad coriónica y finalmente en el polo abembrionario se produce la unión de la decidua, el corion liso y el amnios. Estos tres elementos forman la membrana amniocoriónica o bolsa de las aguas, la cual protruye por el cuello uterino dilatado y se rompe cuando empieza el trabajo de parto, derramando el líquido amniótico. En el polo embrionario, el amnios y el corion ubicados sobre la decidua basal forman la placa coriónica.
Placenta
La placenta posee un componente materno, la decidua basal y un componente fetal, el corion frondoso. Hasta la segunda semana de desarrollo la placenta presenta las lagunas trofoblásticas, etapa lacunar. Desde aquí y hasta el tercer mes, la placenta tiene un desarrollo notable de las vellosidades coriónicas, etapa vellositaria. Por último, desde el cuarto mes en adelante se forman los tabiques deciduales, etapa cotiledónica. En esta etapa, la decidua basal forma tabiques que sobresalen en las cámaras hemáticas o espacios intervellosos (ex lagunas trofoblásticas) pero no logran tomar contacto con la placa coriónica. Estos tabiques delimitan espacios más o menos cúbicos llamados cotiledones, en cuyo interior se encuentran las vellosidades bañadas por la sangre materna. Por el lado materno de la placenta, los cotiledones se evidencian como 15 a 20 áreas poligonales de la decidua basal. Los cotiledones están perforados por las venas endometriales y las arterias espirales de la mucosa uterina. Estas últimas atraviesan la decidua y se abren en los espacios intervellosos depositando sangre que luego es recogida por las venas endometriales. Dentro del cotiledón, se denomina cotiledón fetal a porción vellositaria y cotiledón materno al componente decidual. La presión sanguínea permite que esta sangre oxigenada y con nutrientes tome contacto con las vellosidades coriónicas, se produzca el intercambio y luego sea drenada por las venas endometriales hacia la circulación materna.
Barrera placentaria
Entre la sangre fetal y materna se encuentra un “tabique” de tejido fetal que establece la barrera placentaria. Esta barrera está formada por:
1. El endotelio de los vasos fetales.
2. El mesoderma extraembrionario que rodea a los vasos fetales.
3. El citotrofoblasto.
4. El sincitiotrofoblasto.
La barrera placentaria es bastante permeable y a partir del cuarto mes se adelgaza considerablemente, debido a que se reduce la capa de mesoderma y de citotrofoblasto que la forman. El término “barrera” es inadecuado, ya que muchos fármacos, virus (rubéola, sarampión, poliomielitis), bacterias (Treponema pallidum, causante de la sífilis, y el Toxoplasma gondii, causante de la toxoplasmosis), y moléculas de tamaño pequeño pasan de la sangre materna a la sangre fetal, alcanzando concentraciones suficientes para afectar el desarrollo fetal (Figura 2-12).
Funciones y características de la placenta
Las funciones de la placenta son:
1. Permitir el intercambio de gases entre la sangre fetal y materna: función respiratoria.
2. Posibilitar el aporte de nutrientes de la madre al feto: función nutricia.
3. Eliminar los desechos metabólicos de la sangre fetal: función excretora.
4. Transmitir anticuerpos de la sangre materna al feto, sustancias que atraviesan la barrera placentaria: función inmunológica.
5. Producir hormonas como progesterona, estradiol, gonadotrofina coriónica, somatotrofina: función endocrina.
6. Sintetizar glucógeno, colesterol y ácidos grasos: función metabólica.
La placenta de término tiene forma de disco de 15 a 25 cm. De diámetro tiene 3 cm de espesor y pesa alrededor de 500 g. Después del parto, se separa la decidua basal, quedando en el útero la capa basal y la placenta es expulsada durante el alumbramiento. Al examinar la placenta por el lado materno, se aprecian los relieves que marcan los cotiledones maternos y por el lado fetal se observa la placa coriónica donde se encuentran gruesas arterias y venas, los vasos coriónicos, que convergen hacia el cordón umbilical.
Hacia el final del embarazo la placenta presenta cambios, aumentando el tejido fibroso en el centro de las vellosidades y el depósito de un tejido fibrinoide en las superficies de las vellosidades.
Cordón umbilical
El cordón umbilical mide 50 cm de largo y 2 cm de espesor y se extiende desde el anillo umbilical primitivo, ubicado en la pared abdominal del feto, hasta la inserción en la placa coriónica de la placenta, inserción que suele presentarse en el centro de la superficie fetal de la placenta. El cordón umbilical, cuya cubierta externa corresponde al amnios, contiene dos arterias umbilicales, una vena umbilical, restos del saco vitelino y del alantoides, y entre estos elementos un tejido gelatinoso de origen mesodérmico, la gelatina de Wharton (Figura 2-27).
Amnios
Como hemos visto anteriormente, a partir del tercer mes el amnios y la cavidad amniótica crecen englobando al feto, el cual flota en el líquido amniótico producido por el amnios. Esto tiene una acción de protección, permite los movimientos fetales y evita que se formen adherencias en el feto. El amnios puede ser separado de las membranas fetales y es transparente, firme y avascular; el líquido amniótico es transparente como agua de roca. El líquido es inicialmente producido por las células amniogénicas, pero después del tercer mes, es filtrado de los tejidos placentarios. El volumen del líquido amniótico al final del embarazo es de alrededor de 800 ml y, a partir del quinto mes, el feto deglute volúmenes importantes de él agregándose, consecuentemente, volúmenes de orina fetal al líquido amniótico. El líquido amniótico se recambia cada cuatro horas. También flotan en el líquido células fetales descamadas, que pueden ser aspiradas para su estudio mediante una amniocentesis o punción del saco amniótico (Figuras 2-28a y b).
La existencia de volúmenes bajos de líquido amniótico para cualquier edad gestacional, oligohidroamnios, es indicativo de:
1. Falla placentaria, por disminución del flujo sanguíneo o desprendimiento de la membrana amniocoriónica.
2. Alteración fetal, por agenesia renal u obstrucción de las vías urinarias, donde falta la contribución de la orina fetal al volumen del líquido amniótico.
Por otro lado, los volúmenes altos de líquido amniótico, polihidroamnios o hidroamnios, se presentan cuando el feto no ingiere la cantidad adecuada de este líquido, tal como ocurre en la atresia esofágica o en anomalías graves como la anencefalia.
Saco vitelino y alantoides
El saco vitelino y el alantoides, cuyos restos pueden encontrarse en el cordón umbilical, representan anexos embrionarios que involucionan en la especie humana. El saco vitelino secundario (la yema del huevo de los ovíparos) no tiene en el hombre una función nutritiva. Sin embargo, su importancia radica en que desde la pared del saco vitelino se diferencian y migran hacia las crestas genitales las células germinativas primordiales que darán origen finalmente a los gametos. Además, en la pared del saco vitelino se diferencian los primeros elementos sanguíneos embrionarios. Durante el desarrollo, parte del saco vitelino secundario formará el intestino primitivo y, a medida que el amnios crece y se desarrolla la pared corporal, el saco vitelino se reduce progresivamente, quedando sus vestigios incluidos en el cordón umbilical. El alantoides, pequeño divertículo del saco vitelino que se extiende hacia el pedículo de fijación, si bien carece de función en el hombre, es importante porque los vasos sanguíneos que se desarrollan en su pared se transformarán en los vasos umbilicales. Después del segundo mes, el alantoides involuciona y forma un tubo membranoso, el uraco, conectado con la vejiga urinaria, elemento que está incluido también en el cordón umbilical.
Anexo Figuras
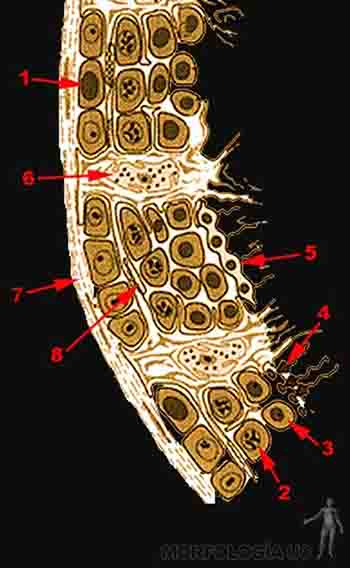
Figura 2-1
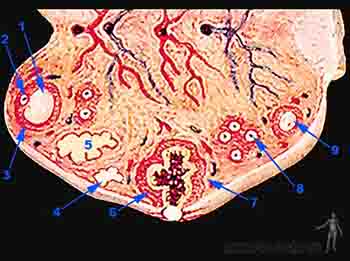
Figura 2-2a
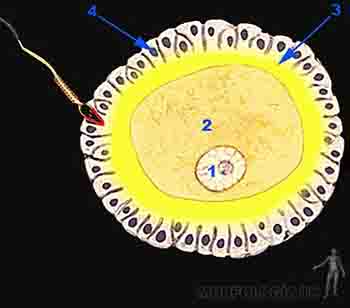
Figura 2-2b
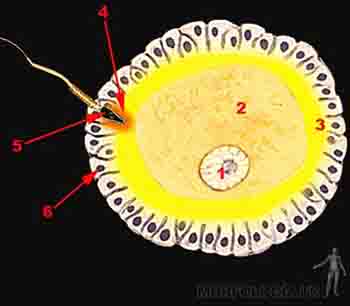
Figura 2-3a
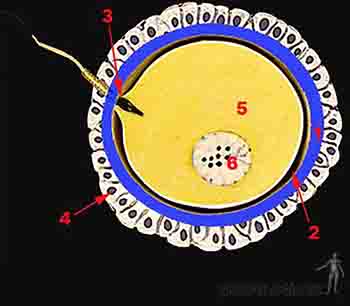
Figura 2-3b
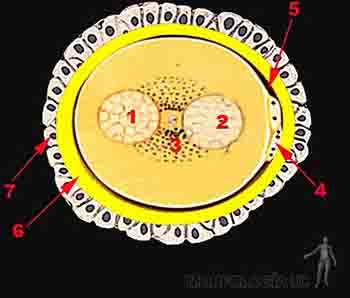
Figura 2-3c
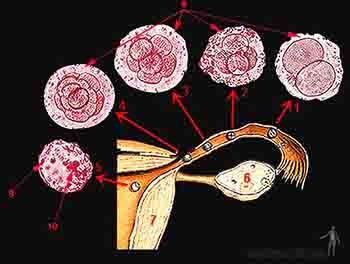
Figura 2-4
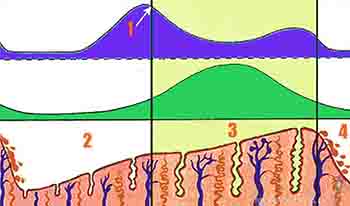
Figura 2-5
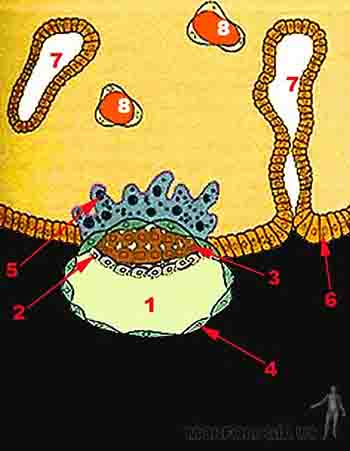
Figura 2-6a
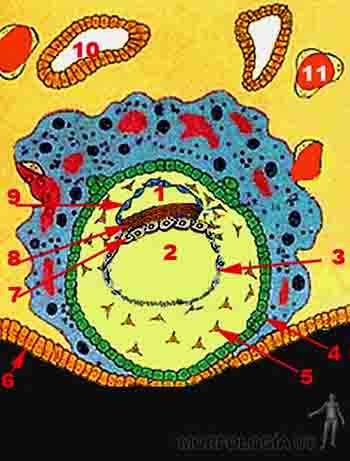
Figura 2-6b
Figura 2-6c
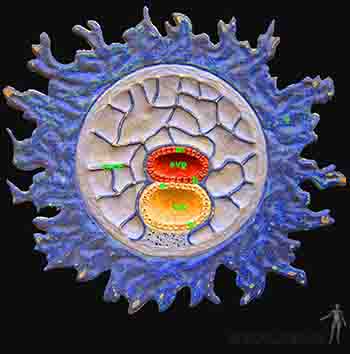
Figura 2-7a
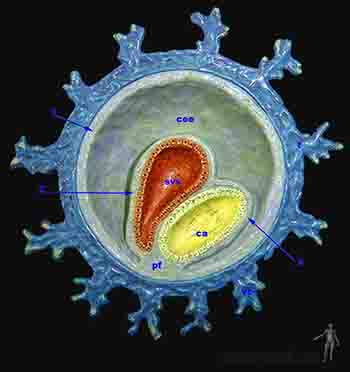
Figura 2-7b
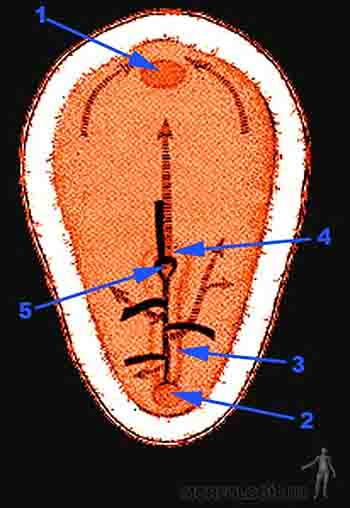
Figura 2-8a
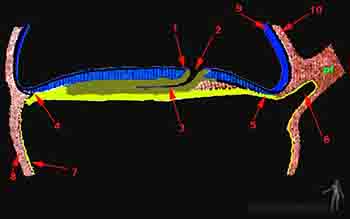
Figura 2-8b
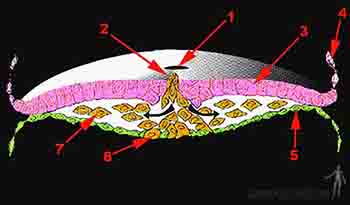
Figura 2-9
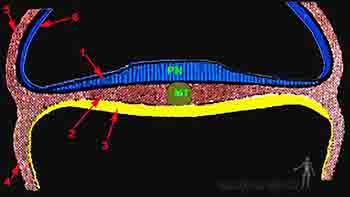
Figura 2-10a
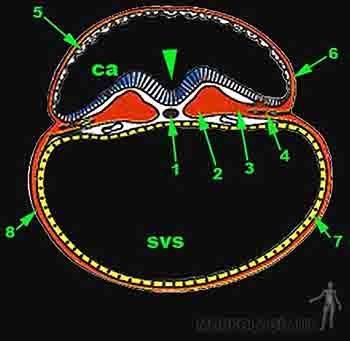
Figura 2-10b
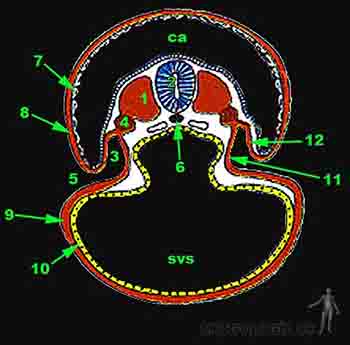
Figura 2-11
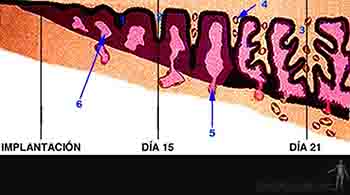
Figura 2-12
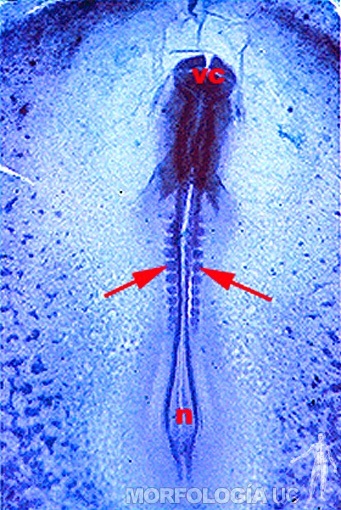
Figura 2-13
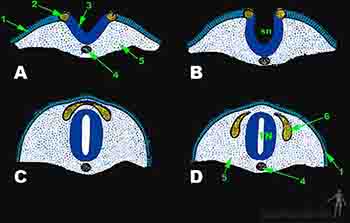
Figura 2-14
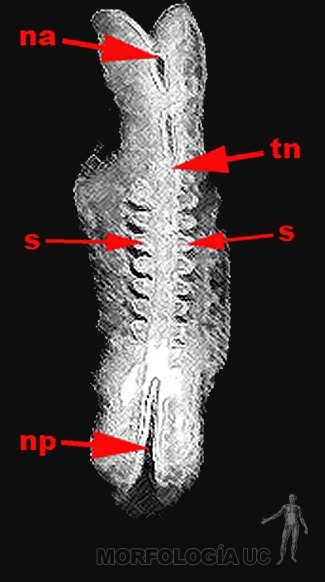
Figura 2-15
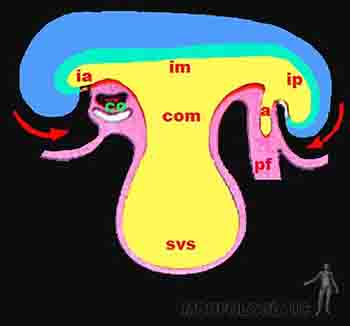
Figura 2-16a
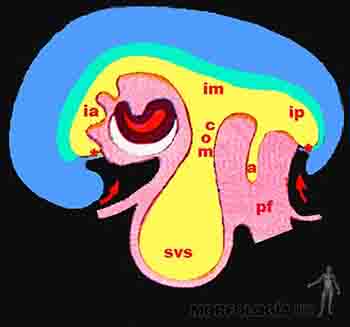
Figura 2-16b
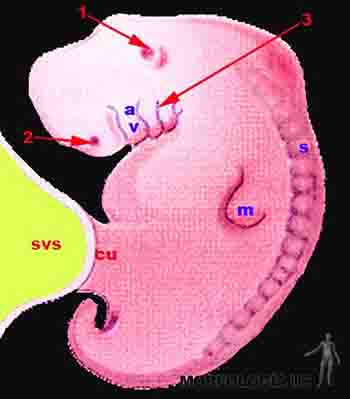
Figura 2-17
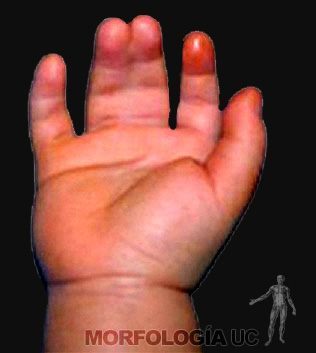
Figura 2-18
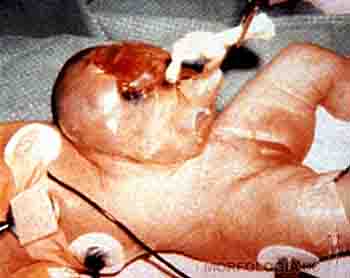
Figura 2-19
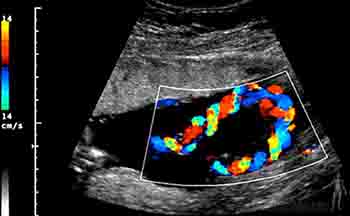
Figura 2-20a
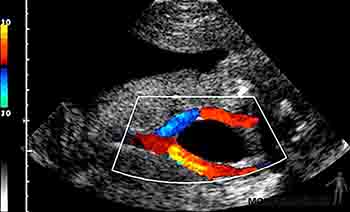
Figura 2-20b
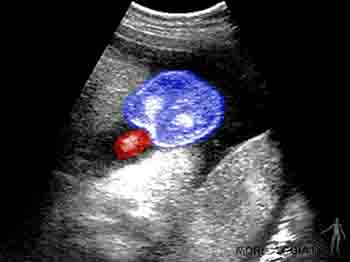
Figura 2-21a
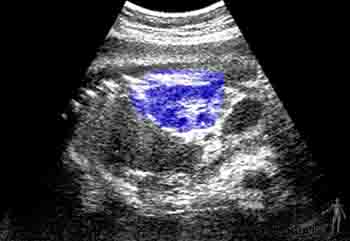
Figura 2-21b
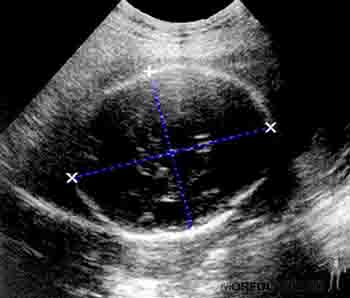
Figura 2-22
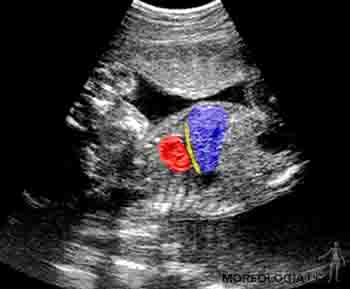
Figura 2-23
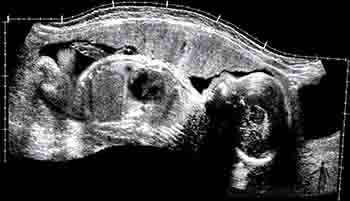
Figura 2-24
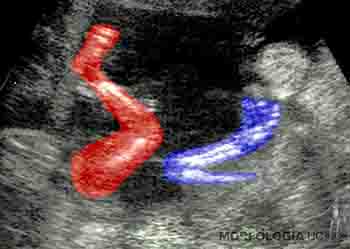
Figura 2-25a
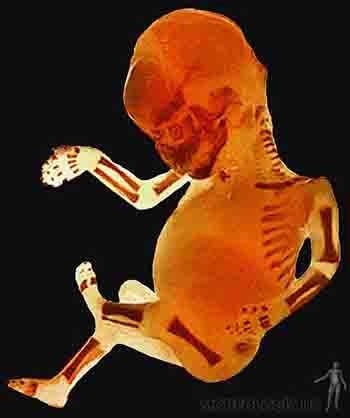
Figura 2-25b
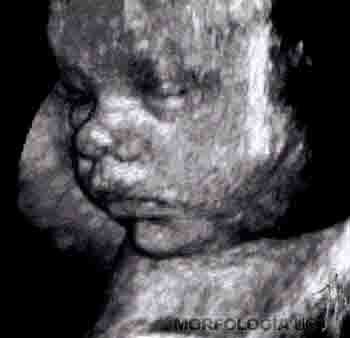
Figura 2-25c
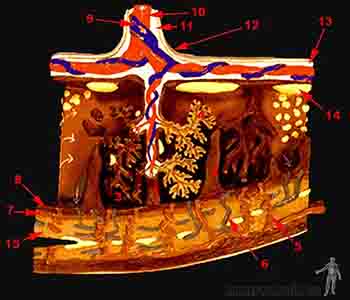
Figura 2-26
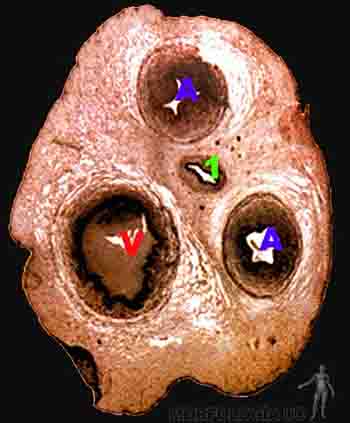
Figura 2-27
Figura 2-28a
Figura 2-28b