Capítulo 3
EL INTESTINO DELGADO
Se suele considerar el intestino impermeable a las macromoléculas.
Sin embargo, tenemos cada día más pruebas de que, incluso en una persona sana, las macromoléculas atraviesan la barrera intestinal, en cantidades insuficientes para tener importancia nutritiva, pero suficientes para desarrollar una actividad antigénica o biológica.
W. A. WALKER y K. J. ISSELBAKER
El intestino, expuesto a estimulaciones antigénicas múltiples, contiene del 70 al 80% de las células productoras de anticuerpos del organismo humano.
WALEED S.W. SHALABY
El intestino delgado merece un estudio detallado, ya que es, a mi parecer, un órgano clave. La mucosa del intestino delgado sirve de barrera entre el medio interior del organismo y los peligrosos factores del medio ambiente: bacterias y alimentos. En la mayoría de las personas, esa barrera desempeña mal su papel y permite que la atraviesen demasiadas macromoléculas. Algunas de estas sustancias son nocivas y su acumulación, junto con factores hereditarios favorables, produce numerosas enfermedades.
A) ARQUITECTURA DEL INTESTINO DELGADO
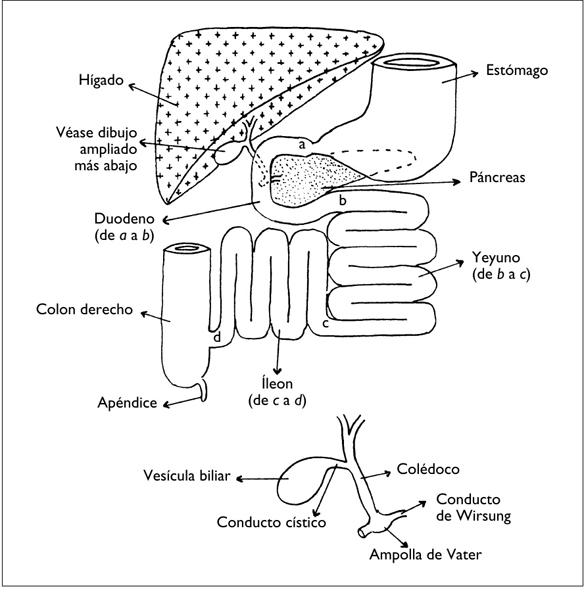
Figura 1. EL INTESTINO DELGADO
1. Anatomía
El intestino delgado es la parte del tracto digestivo comprendida entre el estómago y el colon o, de forma más precisa, entre el píloro y el esfínter ileocecal. Mide de 5 a 6 m y tiene tres partes (figura 1):
• El duodeno, que forma un cuadrado en el cual se sitúa la cabeza del páncreas.
• El yeyuno, que adopta una forma de asas dispuestas horizontalmente.
• El íleon, que está constituido por asas verticales.
Las vías biliares y pancreáticas vierten sus secreciones en el duodeno.
2. Estructura
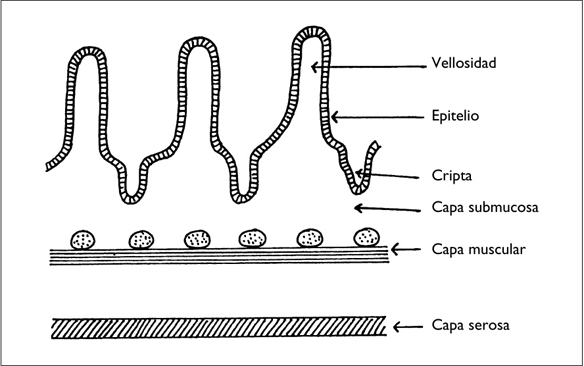
La pared intestinal tiene varias capas que son, empezando por la luz intestinal: la mucosa, la muscular, la submucosa y la serosa (figura 2). Sólo describiremos la mucosa, que desempeña un papel fundamental en la elaboración de mis teorías.
La mucosa es un epitelio formado por una única capa de células que reposan sobre un corion conjuntivo llamado lámina propia. La mucosa presenta numerosas protuberancias, de 0,1 a 0,8 mm de altura, llamadas vellosidades, separadas por depresiones, denominadas criptas. Esta estructura aumenta considerablemente la superficie funcional del intestino, que supera los 100 m2 (Minaire y col., 1990). Si tenemos en cuenta el borde en cepillo de los enterocitos, la superficie puede llegar a los 600 m2.
Figura 2. ESTRUCTURA DE LA PARED DEL INTESTINO DELGADO
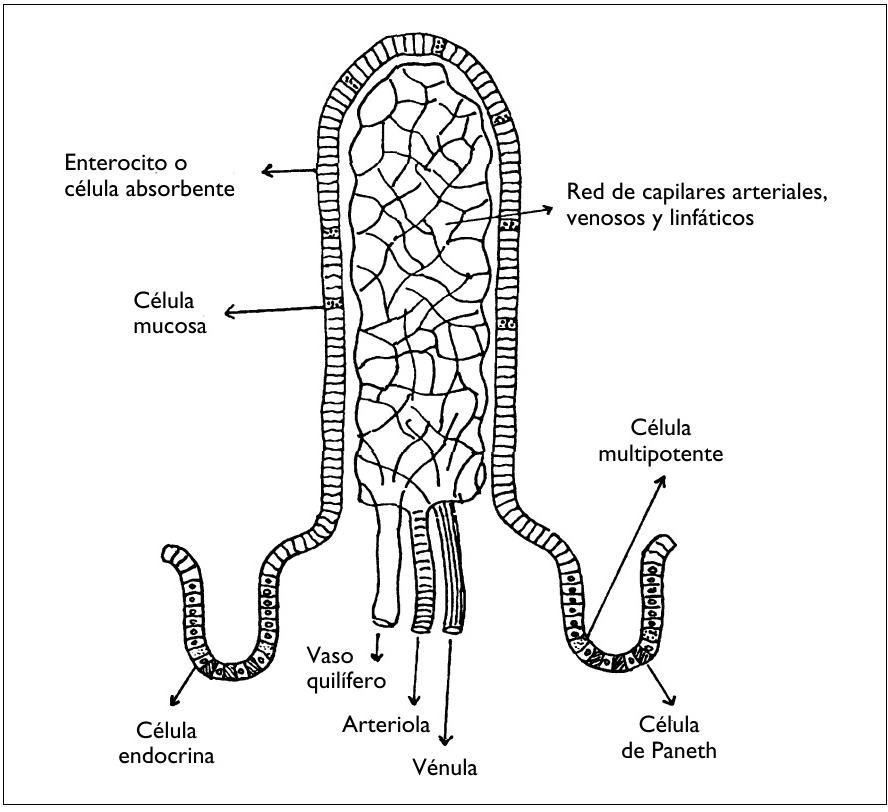
Cada vellosidad tiene una arteriola, una red capilar, una vénula y pequeños vasos linfáticos drenados por un vaso quilífero central (figura 3). También encontramos en la mucosa los folículos linfáticos y las placas de Peyer, de las cuales hablaremos más adelante al describir las defensas del intestino delgado.
B) LA MUCOSA DEL INTESTINO DELGADO
El epitelio está constituido por diversos tipos de células que provienen de una célula de origen pluripotente, situada en las criptas (Haffen y col., 1990). La renovación de las células de las vellosidades se lleva a cabo desde la base hasta la cima. Se distinguen cinco variedades de células:
1. Los enterocitos o células absorbentes
Con diferencia, son las células más numerosas, tienen forma alargada y estrecha, forman una pared (epitelio cilíndrico) interrumpida en algunas zonas por una célula mucosa, y a la altura de las placas de Peyer, por las células M. En el polo apical, los enterocitos tienen un borde en cepillo.
La duración media de vida de los enterocitos es corta: siete días en el caso de los del duodeno y el yeyuno, tres en los del íleon. Las agresiones de elementos intraintestinales acortan su vida, pero la renovación a partir de las células originarias es rápida.
Figura 3. ESTRUCTURA DE UNA VELLOSIDAD Y DE DOS CRIPTAS
Los enterocitos están conectados entre sí por tres tipos de uniones (Minaire y col., 1990):
• Uniones estrechas.
• Uniones intermediarias.
• Desmosomas.
La barrera que forman los enlaces estrechos se manifiesta por la resistencia eléctrica transepitelial.
2. Las células mucosas
Estas células segregan los péptidos trifoliados y el moco.
Los péptidos trifoliados tienen una estructura particular, con tres puntos desulfurados tensos entre seis cisteínas, lo que les confiere un aspecto en hoja de trébol. Su estructura compacta les permite resistir a las enzimas proteolíticas. Probablemente desempeñan un papel importante en el mantenimiento de la integridad de la mucosa intestinal y en la cicatrización de las heridas (Sands y Podolsky, 1996), (Amiranoff, 1996).
Se producen tres litros de moco diariamente, de pH alcalino, cuya función es proteger la mucosa intestinal frente a diversos agresores, como los pH extremos de los jugos digestivos, las enzimas digestivas, las bacterias y los alimentos. El moco es una mezcla de mucinas. Las mucinas son glicoproteínas en las que los glúcidos (glicanos) son mucho más abundantes que los péptidos: el 85% frente al 15%.
Sin embargo, la fracción más interesante está constituida por los péptidos, ya que son extremadamente variados. Existen centenares de mucinas diferentes. Gum y col. (1989) demostraron que los genes de las mucinas están reagrupados en dos zonas, una situada en el cromosoma 11ylaotraenelcromosoma7.
El notable polimorfismo de las mucinas tiene un significado biológico. Porchet y col. (1991) plantearon la hipótesis de que cada mucina estaba especializada en la neutralización de un agresor determinado.
3. Las células de Paneth
Están situadas en el fondo de las criptas. Estas células secretan las lisozimas y las defensinas (Ganz, 2000).
4. Las células endocrinas
Están localizadas principalmente en el fondo de las criptas. Segregan varios péptidos, los más importantes son: el GIP (gastric inhibitory peptide), la secretina, la colecistokinina, el péptido YY, el GLP-1 (glucagon like peptide 1) y el enteroglucagón.
5. Las células M
Representan entre el 5 y el 10% del total. Las estudiaremos junto con las defensas inmunitarias del intestino delgado.
C) FUNCIÓN DEL INTESTINO DELGADO
Aparte de su función en las reacciones de inmunidad y de tolerancia, que examinaremos más adelante, el intestino delgado desempeña tres funciones:
1. Participa en la digestión de los alimentos
La digestión consiste en la degradación de grandes moléculas complejas en pequeñas moléculas simples. En una situación ideal:
• Los polisacáridos son degradados en azúcares simples.
• Los lípidos son degradados en cuerpos grasos simples.
• Las proteínas son degradadas en aminoácidos.
La digestión se realiza gracias a las enzimas glicolíticas, lipolíticas y proteolíticas presentes en las diversas secreciones que se vierten en el tubo digestivo: saliva, jugo gástrico, bilis y, principalmente, jugo pancreático y jugo intestinal.
La digestión se efectúa en el duodeno y termina en el yeyuno y el íleon.
2. Asegura una absorción selectiva de las sustancias digeridas
Esta absorción se efectúa en las vellosidades, mientras que, de forma inversa, en las criptas se produce una secreción. Los productos de la digestión pueden atravesar la barrera intestinal por dos vías (Minaire y col., 1990):
a) Entre los enterocitos, es decir, en forma de un transporte pasivo cuyo obstáculo principal son las uniones estrechas;
b) A través de los enterocitos, atravesando primero la membrana apical y después la membrana basolateral. Se trata de un transporte activo con pinocitosis o endocitosis por parte del receptor, que obtiene la energía del adenosintrifosfato (ATP).
La absorción de las diversas sustancias está regulada. Así, la absorción de hierro, por ejemplo, debe ser suficiente, pero no demasiado elevada, pues se correría el riesgo de contraer una grave enfermedad: la hemocromatosis.
Los productos de la digestión de los glúcidos y de las proteínas, así como de los triglicéridos de cadena media, se drenan a través de la sangre. Estos últimos pasan, por tanto, por el hígado. Los productos de la digestión de los lípidos se drenan a través de la linfa.
3. Permite la progresión del quilo
Las sustancias que se digieren en la luz intestinal constituyen el quilo. El quilo avanza en el interior del intestino delgado y pasa al colon. Este tránsito es provocado por las contracciones de los músculos lisos de la pared del intestino delgado, que producen un movimiento peristáltico que hace progresar la ración alimenticia.
D) LA FLORA BACTERIANA DEL INTESTINO DELGADO
El tracto digestivo del hombre contiene alrededor de 1014 bacterias, lo que equivale más o menos a diez veces el número total de células del organismo (Cerf, 1991). Desde el nacimiento, las bacterias aportadas por el aire y por los alimentos invaden el tubo digestivo.
La flora varía en cantidad según las zonas:
• Tiene una abundancia media en la boca.
• Es más escasa en el estómago, ya que la acidez gástrica destruye el 99% de los gérmenes.
• Es más numerosa en el intestino delgado superior, donde se estima el número de bacterias por mililitro entre 103 y106, según los autores.
• Es más abundante aún en el intestino delgado inferior.
• Es extremadamente abundante en el colon, hasta el punto de que las bacterias representan más del 50% del peso de las heces.
La composición de la flora varía según el nivel del intestino. Los microorganismos aerobios, presentes en la parte superior del tubo digestivo, son progresivamente sustituidos por microorganismos anaerobios, que constituyen el 99% del total de la flora en el colon. En el intestino delgado se da una confluencia de las dos variedades de flora:
• En el duodeno y el yeyuno encontramos principalmente bacterias aerobias, en particular colibacilos, enterococos, estreptococos, estafilococos, Pseudomonas, Enterobacter, Citrobacter y Klebsiella (Cerf, 1991). No hay bacterias anaerobias estrictas y pocos lactobacilos.
• En el íleon, por el contrario, predominan las anaerobias.
En total, alrededor de 500 especies bacterianas cohabitan en el intestino delgado (Floch y Moussa, 1998). Se distingue una flora de paso, incapaz de desarrollarse, y una flora residente, capaz de multiplicarse y de implantarse. Esta última se encuentra sobre todo en el intestino delgado terminal y principalmente en el colon.
En la composición de esta flora intestinal influyen varios factores:
1) La acidez gástrica: en caso de aclorhidria o de hipoclorhidria, el número de bacterias se multiplica por 10.000 en el intestino delgado superior, aumentan los lactobacilos y aparecen bacterias anaerobias.
2) El peristaltismo, principalmente activo en el intestino delgado superior, tiende a empujar los gérmenes hacia el extremo inferior del tubo digestivo.
3) Las interacciones entre bacterias: algunas especies inhiben o facilitan el desarrollo de otras especies (Ducluzeau, 1993).
4) La alimentación: está demostrado que un régimen rico en carne favorece el desarrollo de una flora de «putrefacción», mientras que un régimen vegetariano induce la proliferación de una flora de «fermentación». Veremos más adelante que la dietética que preconizo modifica la flora intestinal.
5) Los anticuerpos secretados por los plasmocitos de la pared intestinal. Los más importantes son los IgA secretores que inhiben la proliferación microbiana e impiden la adherencia de bacterias al epitelio de la mucosa.
6) La mucosidad, que obstaculiza la acción de algunas bacterias y que, a la inversa, protege a otras contra los efectos destructores del ácido clorhídrico, de los anticuerpos y de las enzimas digestivas.
7) La capacidad de algunas bacterias de adherirse a los receptores específicos de las células epiteliales, lo que puede alterar la morfología y las funciones de las vellosidades.
Es curioso comprobar que pocos investigadores se han interesado por esta inmensa población bacteriana que habita en nuestro intestino. Asimismo, nuestros conocimientos son limitados sobre este tema. Se distinguen de forma habitual dos estados:
• Un estado fisiológico en el cual la flora es saprofita, ocupa la luz intestinal y es beneficiosa. Se establece una simbiosis entre los gérmenes y el huésped humano. Las bacterias se alimentan de nutrientes, es decir, de los productos que resultan de la digestión de los alimentos. Viven a expensas de su huésped, pero ejercen acciones beneficiosas: completan la digestión de algunos alimentos, degradan los pigmentos biliares, participan en la fabricación de la vitamina K, frenan el desarrollo de las levaduras y los hongos (Besson, 1994b) y liberan poliaminas que, en dosis fisiológicas, son nutritivas para los enterocitos (Loser y col., 1999).
• Un estado patológico en el cual la flora se vuelve patógena: una bacteria peligrosa prolifera en exceso y provoca una enfermedad, ya sea al liberar una toxina (colibacilos, estafilococos), o bien al dañar, incluso atravesar, el epitelio de la mucosa (Shigella, Salmonella) (Cerf, 1993).
En mi opinión, las bacterias intestinales, y más concretamente las aerobias del intestino delgado superior, son las responsables de muchas otras enfermedades. Podemos implicar a Klebsiella en el desarrollo de la espondilitis anquilosante, a Proteus mirabilis en la poliartritis reumatoidea, a Yersinia enterocolitica en la hipertiroiditis de Basedow, etc.
La mayoría de las bacterias que se encuentran en el intestino delgado están muertas, lo que facilita su descomposición en péptidos, lipopolisacáridos y otras sustancias más o menos peligrosas.
Sería muy útil que se efectuasen investigaciones más exhaustivas sobre las relaciones entre bacterias intestinales y patología. Dichas investigaciones podrían inspirarse en trabajos (Megraud, 1993) que han demostrado claramente que Helicobacter pylori, una bacteria de la mucosa gástrica, es el mayor agente causal de la úlcera gastroduodenal y de algunos cánceres gástricos.
E) LAS DEFENSAS DEL INTESTINO DELGADO
La mucosa del intestino delgado es un inmenso filtro cuya superficie funcional llega a ocupar 100 m2,incluso600m2 si se consideran los bordes en cepillo, cuyo espesor es muy reducido (0,025 mm), ya que los enterocitos están dispuestos en una sola capa. Sin embargo, este epitelio delgado es la única barrera que separa nuestro medio interior de ciertos agentes nocivos del medio ambiente: parásitos, bacterias, virus y alimentos digeridos de forma incompleta. La mucosa del intestino delgado está protegida por dos sistemas de defensas: no inmunes e inmunes.
1. Defensas no inmunes
Intervienen en primera línea y comprenden (Sarker y Gyr, 1992):
• El jugo gástrico, que, por su acidez, tiene propiedades bactericidas.
• El jugo pancreático y la bilis, que fraccionan las macromoléculas alimentarias y forman una corriente líquida que arrastra numerosos gérmenes.
• La motricidad del intestino producida por el peristaltismo.
• La rápida renovación de las células epiteliales de la mucosa.
• La flora intestinal, ya que los gérmenes saprofitos impiden la multiplicación de los gérmenes patógenos.
• Las secreciones intestinales de las criptas.
• La mucosidad de los péptidos trifoliados y numerosas variedades de mucinas, cada una de las cuales puede estar especializada en la neutralización de un agresor (Porchet y col., 1991).
• La lisozima, una enzima que elimina muchas bacterias de las paredes (Ganz, 2000).
• Las defensinas, que son péptidos antimicrobianos que se sintetizan a un ritmo mucho más rápido que la velocidad de multiplicación de las bacterias (Nicolas y col., 1992).
2. Defensas inmunes
Actualmente se conocen bastante bien (Cerf-Bensussan y col., 1991) y se apoyan en dos tipos de estructuras:
a) Células diseminadas en la mucosa
1) Los linfocitos B y los plasmocitos segregan mucha más IgA que IgM e IgG. La situación es muy diferente de la de la sangre, en la que las proporciones son inversas: hay más cantidad de IgG, menor de IgM y mucha menos de IgA. Por otra parte, las IgA sanguíneas son monómeras, mientras que las IgA intestinales son dímeras: las dos unidades están unidas por la cadena J y por la pieza secretora (figura 4).
2) Los linfocitos T, de dos clases: auxiliares de tipo CD4 y citotóxicos de tipo CD8.
3) Los macrófagos son numerosos, mientras que los mastocitos, los neutrófilos polinucleares y los eosinófilos polinucleares son raros.
b) Los folículos linfáticos o placas de Peyer
Son formaciones complejas recubiertas por un epitelio de superficie especializada. Este epitelio, que carece de vellosidades, contiene unas células particulares, las células M, que se intercalan entre los enterocitos (Owen, 1994) (figura 5).
Las células M son muy pobres en lisosomas, están desprovistas del borde en cepillo, emiten largos pseudópodos y no sintetizan IgA.
Estas cuatro propiedades, junto a la ausencia de células mucosas en las placas de Peyer, explican por qué la mayoría de los antígenos que proceden de la luz intestinal penetran en las células M antes que en los enterocitos.
Figura 4. ESTRUCTURA DE UNA IgA SECRETORA
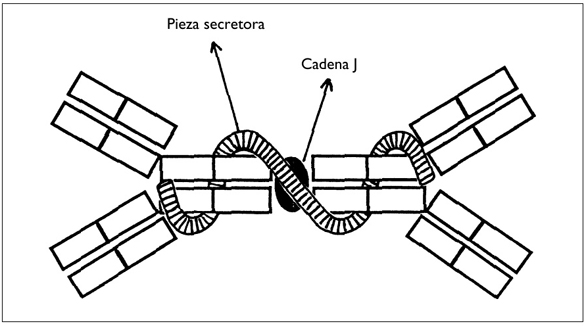
Figura 5. LA CÉLULA M
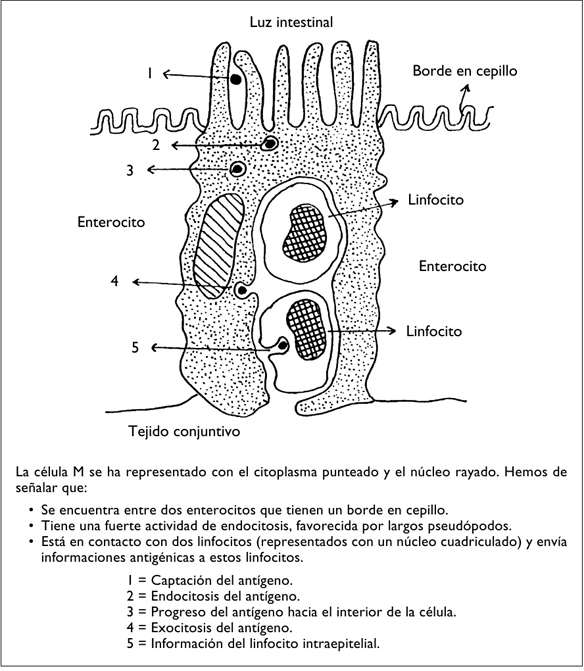
El citoplasma de las células M forma unos repliegues donde se alojan los linfocitos T y B, los plasmocitos y los macrófagos. Es importante señalar que la membrana basal es discontinua en los lugares ocupados por las células M, lo que facilita todavía más este alojamiento. Las células captan los antígenos procedentes del medio y los presentan a los linfocitos, por lo que se las denomina CPA (células presentadoras de antígenos).
F) LA TOLERANCIA ORAL
1. Necesidad de la tolerancia oral
La mucosa del intestino delgado constituye una barrera protectora contra los heteroantígenos, pero no es una barrera perfecta. En una persona normal, las macromoléculas (lipopolisacáridos, péptidos y también proteínas) atraviesan la pared intestinal (Tome, 1990). Lo hacen en una proporción relativamente baja, pero suficiente para desencadenar una respuesta inmunitaria.
En los niños se observan a menudo respuestas de ese carácter, que se reflejan en la intolerancia a la leche de vaca o en la presencia sistemática de anticuerpos contra la albúmina bovina en caso de diabetes juvenil reciente (Karjalainen y col., 1992). Estos fenómenos se pueden atribuir a una permeabilidad excesiva del intestino inmaduro, si bien algunos heteroantígenos llegan directamente a través de la circulación sanguínea.
En un adulto con buena salud no ocurre lo mismo, y es raro detectar anticuerpos contra las bacterias intestinales o las proteínas alimenticias, u observar accidentes de alergia digestiva. Por tanto, se puede suponer que el intestino delgado induce una respuesta de tolerancia respecto a los heteroantígenos con los cuales entra en contacto. Ese proceso se denomina tolerancia oral.
2. Demostración de la tolerancia oral
En el transcurso de los últimos años se han publicado diversos trabajos que confirman la realidad de la tolerancia oral (Marx, 1991). Diversos autores han utilizado modelos animales experimentales de enfermedades autoinmunes y han comprobado que un mismo antígeno tiene distintos efectos según su vía de entrada.
• Administrado inyectado en un vaso sanguíneo, el antígeno provoca el desarrollo de una afección autoinmune.
• Administrado por vía oral, el antígeno puede prevenir la aparición de un estado autoinmune o bloquear su evolución si se ha desarrollado. Este efecto beneficioso no se ha comprobado en todos los animales, pero sí en algunos.
El fenómeno se ha observado en varios modelos:
• La encefalomielitis alérgica experimental (EAE) en la rata, equivalente a la esclerosis múltiple humana, cuyo antígeno causante es la proteína básica de la mielina.
• La uveítis de la rata, inflamación ocular producida por la sustancia retiniana S.
• La artritis del colágeno en varios animales, parecida a la poliartritis reumatoidea humana, cuyo antígeno causal es el colágeno de tipo II.
3. Mecanismos de la tolerancia oral
Todavía no se conocen bien los mecanismos de la tolerancia oral, pero es muy probable que activen ciertas funciones de las células M y de los linfocitos T, característicos de la pared del intestino delgado.
G) LA HIPERPERMEABILIDAD DEL INTESTINO DELGADO
1. Aspecto fisiológico
Hace algunas décadas se creía que la absorción selectiva del intestino delgado era perfectamente eficaz y que sólo dejaba filtrar el agua, los iones, las vitaminas, los azúcares simples, las grasas simples y los aminoácidos.
En la actualidad se sabe que, incluso en una persona sana, la impermeabilidad del intestino delgado es imperfecta. Fairclough y col. (1980) demostraron que tanto los péptidos pequeños como los aminoácidos franquean fácilmente la barrera intestinal. Las moléculas más voluminosas, en particular los péptidos más grandes y algunas proteínas, atraviesan la mucosa (Tome, 1990) en menor proporción, aunque no insignificante. En individuos sanos, las proteínas del huevo y de la leche de vaca se han detectado en la sangre pocas horas después de comer (Foucard y col., 1975) (Husby y col., 1985). Walker e Isselbaker (1974) estiman que alrededor de 1/1.000 de las proteínas intactas llegan a la sangre portal.
La permeabilidad del intestino delgado está controlada por los péptidos reguladores, sintetizados unos por el sistema nervioso central o periférico, y otros procedentes de células endocrinas diseminadas por el tubo digestivo y el páncreas (Brown, 1993).
2. Aspecto patológico
Es necesario un paso exagerado de proteínas alimenticias para explicar:
• Los accidentes de atopia después del consumo de leche o huevos (Paganell y col., 1979).
• La intolerancia a la leche de vaca en el adulto.
• La intolerancia al gluten en el adulto.
• Las migrañas debidas a la leche, al trigo o a los huevos, que se curan con la supresión del alimento causal.
En el caso de muchas enfermedades estudiadas en este libro, se ha demostrado que había una mayor permeabilidad del intestino delgado en la mayoría o la totalidad de los pacientes. Ampliaré esta idea al tratar la poliartritis reumatoidea, la espondilitis anquilosante, la diabetes insulinodependiente, la nefropatía por IgA y la enfermedad de Crohn, entre otras.
3. Métodos para medir la permeabilidad del intestino delgado
Se utilizan sustancias inertes que traspasan la mucosa sin transformación ni ayuda, y que después se eliminan por la orina, en la que se mide su concentración tras la carga oral (Heresbach y col., 1994) (Bjarnason y col., 1995).
Los principales productos utilizados son:
• Lactulosa/manitol.
• Cromo 51/EDTA.
• L-Ramnosa.
• Polietilenglicol.
• Tecnecio 99/EDTA.
4. Causas de la hiperpermeabilidad
La vía transcelular es sólida y en pocas ocasiones se ve alterada. Sin embargo, una agresión de los enterocitos puede provocar la muerte de algunas células y el acortamiento y la atrofia de las vellosidades. Es cierto que el epitelio se reconstituye a partir de las criptas, pero los enterocitos jóvenes son pobres en enzimas y menos eficaces.
La vía intracelular es mucho más frágil. El talón de Aquiles de la mucosa son los pliegues circulares o las válvulas conniventes que unen los enterocitos y que pueden distenderse.
En las personas predispuestas por ciertos factores genéticos (enzimas, mucinas), diversos elementos pueden determinar un aumento de la permeabilidad:
a) Algunas bacterias
Algunos microorganismos pueden multiplicarse en exceso, adherirse a las células epiteliales y lesionarlas de una manera más o menos grave, liberar toxinas y provocar lesiones inflamatorias importantes (Cerf, 1993).
Entre las bacterias peligrosas destacan los estafilococos, estreptococos, colibacilos, y los géneros Klebsiella, Shigella, Yersinia, Salmonella, Campylobacter, Aeromonas y Clostridium. Se han descubierto dos bacterias que viven en el mucus intestinal: Helicobacter cinaedi y Helicobacter fenneliae. Dichas bacterias merecerían investigaciones más profundas.
b) Algunos alimentos
Examinaremos el terrible impacto de la alimentación moderna en el siguiente capítulo.
c) Algunos medicamentos
Los antiinflamatorios no esteroideos (AINE), los salicilatos y los corticoides tienen efectos nocivos sobre la pared del intestino delgado. Dichos medicamentos actúan a través de varias vías (Gargot y Chaussade, 1993) (Scheiman, 1996):
• Separan los enterocitos, lo que aumenta la permeabilidad intestinal.
• Disminuyen localmente la producción de prostaglandinas, lo que ocasiona una disminución de la mucina, de los fosfolípidos de superficie y de los bicarbonatos, y provoca una merma de los medios de defensa de la mucosa.
• Suelen inducir una inflamación, observada por la presencia de numerosos granulocitos en las heces y por el aspecto de la mucosa mediante exámenes endoscópicos. Dicha inflamación daña a veces los vasos pequeños y provoca una ulceración.
• Inhiben las mitocondrias y la ciclooxigenasa, frenando así las divisiones de los enterocitos.
Estos medicamentos actúan como armas de doble filo cuando se emplean para tratar las enfermedades inflamatorias relacionadas con el paso de macromoléculas bacterianas o alimentarias a través de la barrera intestinal: por una parte, calman momentáneamente la inflamación y alivian los dolores de los enfermos, pero, por otra parte, aumentan la permeabilidad del intestino delgado, permitiendo que lo atraviesen nuevas macromoléculas que perpetúan la enfermedad.
d) El interferón gamma
Adams y col. (1993) pusieron en evidencia los efectos de esta citocina. La permeabilidad intestinal normal se traduce por una resistencia eléctrica transepitelial superior a 1.000 ohms por centímetro cuadrado. Después de una exposición al interferón gamma, esa resistencia se reduce de forma neta.
El interferón gamma se fija a los receptores de membrana del polo basal de los enterocitos. Una exposición de un día a la citocina ocasiona una disminución de la resistencia eléctrica durante cinco días, seguida de un retorno a la normalidad. El interferón gamma no altera los enterocitos y no modifica los pasos transcelulares, sino que amplía los pliegues circulares entre los enterocitos, lo cual aumenta la permeabilidad intercelular.
Otra citocina, la interleucina 4, produce los mismos efectos que el interferón gamma (Colgan y col., 1994).
5. Consecuencias de la hiperpermeabilidad
Los residuos bacterianos y alimentarios traspasan en cantidad excesiva la pared del intestino delgado y penetran en la circulación general. Estas macromoléculas, junto con factores genéticos, son, desde mi punto de vista, las causantes de numerosas enfermedades.
6. Algunas palabras sobre el colon derecho
El colon derecho (colon ascendente) es la parte inicial del colon comprendido entre el esfínter ileocecal y el ángulo cólico derecho, de donde parte el colon transverso.
El colon derecho contiene una flora bacteriana abundante y variada. Se trata clásicamente de bacterias anaerobias, aunque restos de bacterias aerobias, en particular péptidos, pueden llegar hasta el colon. En la actualidad, está demostrado que la pared del colon absorbe ciertas sustancias resultantes del metabolismo de esas bacterias.
Por otra parte, algunas moléculas escapan de la digestión en el intestino delgado: almidón, polisacáridos o proteínas. Dichos alimentos pueden ser digeridos en el colon por las bacterias anaerobias. Entre el 5 y el 10% de los nutrientes se absorben en el colon.
Una hiperpermeabilidad de la pared del colon derecho tiene, con toda probabilidad, consecuencias nefastas.