2
El nacimiento de lo bueno y lo malo
La naturaleza ha situado a la humanidad bajo el gobierno de dos amos soberanos, el dolor y el placer.
—Jeremy Bentham, Introducción a los principios de la moral y la legislación.
A primera vista, la diversidad del reino animal parece extraordinaria; desde las hormigas a los cocodrilos, desde las abejas a los babuinos y desde los crustáceos a los gatos, las variaciones de los animales parecen innumerables. No obstante, si investigáramos un poco más, también podríamos concluir fácilmente que lo que resulta extraordinario sobre el reino animal es cuán poca diversidad existe en realidad. Casi todos los animales de la Tierra poseen el mismo plan corporal. Todos cuentan con un frente donde se aloja la boca, el cerebro y los principales órganos sensoriales (como los ojos y orejas), y todos tienen una parte trasera por donde expulsan sus desechos.
Los biólogos evolutivos denominan a los animales que poseen ese plan corporal «bilaterales», ya que cuentan con una simetría bilateral. Esto contrasta con nuestros primos animales más lejanos —pólipos coralinos, anémonas y medusas—, que presentan un plan corporal de simetría radial; es decir, cuentan con partes similares que se disponen alrededor de un axis central, sin frente ni parte trasera. La diferencia más evidente entre estas dos categorías es cómo se alimentan. Los bilaterales introducen comida en sus bocas y luego defecan los desechos por sus traseros. Los animales radialmente simétricos solo tienen una apertura —una «boca-trasero», por llamarla de alguna manera— que lleva el alimento al estómago y luego lo escupe. Los bilaterales son, sin lugar a dudas, los más educados de los dos.

Figura 2.1
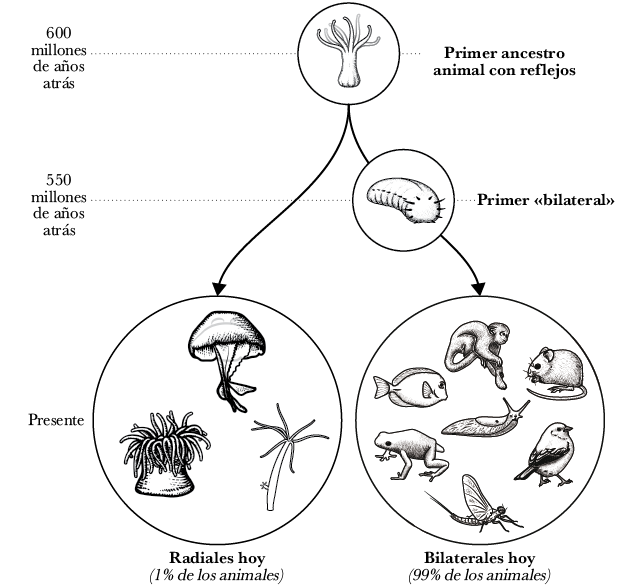
Figura 2.2
Se cree que los primeros animales han sido radialmente simétricos y, sin embargo, hoy en día la mayoría de las especies animales son bilateralmente simétricas. A pesar de la diversidad de los bilaterales modernos —desde los gusanos a los seres humanos—, todos descienden de un único ancestro bilateral en común que vivió alrededor de 550 millones de años atrás. ¿Por qué, dentro de este único linaje de animales antiguos, los planes corporales cambiaron de la simetría radial a la simetría bilateral?
Los planes corporales radialmente simétricos funcionan bien con la estrategia coralina de esperar por el alimento, pero funcionan de manera terrible con la estrategia de caza de desplazarse hacia el alimento. Los planes corporales radialmente simétricos, si se movieran, requerirían que el animal tuviera mecanismos sensoriales para detectar la ubicación del alimento en cualquier dirección, y que luego tuviera la maquinaria para trasladarse hacia él. En otras palabras, tendrían que tener la capacidad simultánea de detectar y moverse en todas las direcciones. Los cuerpos bilateralmente simétricos hacen que el movimiento sea mucho más simple. En lugar de necesitar un sistema motor para trasladarse en cualquier dirección, solo necesitan un sistema motor que se traslade hacia adelante y uno que gire. Los cuerpos bilateralmente simétricos no necesitan escoger la dirección exacta; deben escoger si ajustar el movimiento hacia la derecha o hacia la izquierda.

Figura 2.3: ¿Por qué la simetría bilateral es mejor para la navegación?
Ni siquiera los ingenieros modernos han encontrado aún una mejor estructura para el desplazamiento. Los coches, los aviones, los barcos, los submarinos y casi toda la maquinaria de desplazamiento construida por seres humanos son bilateralmente simétricos. Es, simplemente, el diseño más eficiente para un sistema de movimiento. La simetría bilateral permite que un aparato de movimiento esté optimizado para tomar una sola dirección (hacia adelante) mientras que resuelve el problema del desplazamiento con la añadidura de un mecanismo para girar.
Hay una observación adicional sobre los bilaterales, quizás la más importante: son los únicos animales que poseen cerebro. Esto no es una coincidencia. El primer cerebro y el primer cuerpo bilateral comparten el mismo propósito evolutivo inicial: permitir que los animales se desplacen con control de dirección. La direccionalidad fue el avance #1.
Desplazamiento por direccionalidad
Aunque no sabemos exactamente cómo lucían los primeros bilaterales, los fósiles indican que eran criaturas sin patas del tamaño de un grano de arroz 52. Los restos indican que aparecieron por primera vez en algún punto del periodo Edicárdico, una era que tuvo lugar de 635 a 539 millones de años atrás. El fondo marino durante este periodo estaba repleto de esteras microbianas, verdes y viscosas, en sus áreas poco profundas, y formaban vastas colonias de cianobacterias que se exponían a la luz solar. Los animales pluricelulares capaces de percibir su entorno, como los corales, las esponjas de mar y las primeras plantas, habrían sido muy comunes.
Se cree que los nematodos modernos no han cambiado demasiado desde los primeros bilaterales; estas criaturas nos ofrecen una ventana al funcionamiento interno de nuestros ancestros semejantes a los gusanos. Los nematodos son, casi literalmente, solo una plantilla básica de un bilateral: no mucho más que una cabeza, una boca, un estómago, un trasero, algunos músculos y un cerebro.
Figura 2.4: El mundo edicárdico
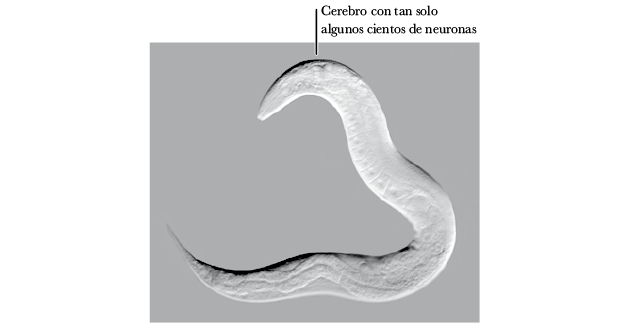
Figura 2.5: El nematodo C. elegans
Los primeros cerebros eran, al igual que los de los nematodos, casi con toda seguridad muy simples 53. El nematodo más estudiado, el Caenorhabditis elegans, posee tan solo 302 neuronas, un número minúsculo en comparación con los 85 mil millones de los seres humanos 54. No obstante, los nematodos exhiben un comportamiento increíblemente sofisticado, a pesar de su cerebro minúsculo. Lo que hace un nematodo con su irremediable cerebro simple nos brinda información sobre lo que los primeros bilaterales hacían con los suyos.
La diferencia más evidente del comportamiento entre los nematodos y animales más antiguos, como los corales, es que los nematodos pasaban mucho tiempo moviéndose. Propongo este experimento: dejen a un nematodo a un costado de una placa de Petri y coloquen un trozo diminuto de comida al otro lado. Observaríamos tres cuestiones: primero, que siempre encuentra la comida 55. Segundo, que la encuentra con mucha más rapidez que si estuviera moviéndose al azar. Y tercero, que no se traslada directamente hacia el alimento, sino que lo hace con un movimiento circular 56.
El gusano no utiliza la visión; los nematodos no pueden ver. No tienen ojos para proyectar una imagen útil para desplazarse. En cambio, el gusano utiliza el olfato. Cuanto más se acerca a la fuente de un olor, mayor es la concentración de ese olor. Los gusanos se valen de esta estrategia para encontrar alimento. Lo único que debe hacer es girar hacia la dirección donde la concentración de partículas de alimento se encuentra en aumento y alejarse de la dirección en la que esa concentración está decreciendo. Resulta muy elegante lo simple y a la vez efectiva que es esta estrategia de desplazamiento. Puede resumirse con dos reglas:
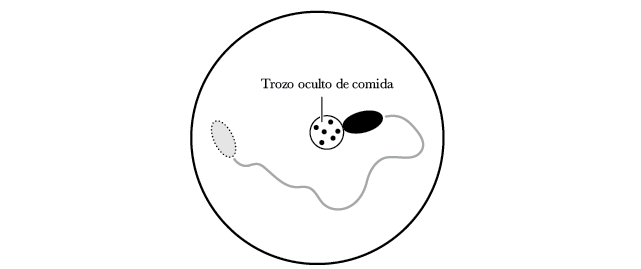
Figura 2.6: Nematodo desplazándose hacia la comida
- Si el olor del alimento se intensifica, sigue hacia adelante.
- Si el olor del alimento decrece, gira.
Este fue el avance de la «direccionalidad». Resulta que, para trasladarse con éxito en el mundo complejo del fondo del océano, no se necesita un entendimiento de ese mundo de dos dimensiones. No es necesario saber dónde te encuentras, dónde está el alimento, qué caminos deberías tomar, cuánto tiempo te llevaría hacerlo, o realmente nada significativo sobre el mundo. Lo único que necesitas es un cerebro que conduzca un cuerpo bilateral hacia olores más intensos y lo aleje de los menos intensos.
La direccionalidad puede utilizarse no solo para movilizarse hacia algo, sino también para alejarse. Los nematodos poseen células sensoriales que detectan la luz, la temperatura y el tacto. Se alejan de la luz, donde los depredadores pueden verlos con facilidad; se alejan del calor y el frío nocivos, donde las funciones corporales se vuelven más difíciles de llevar a cabo, y se alejan de superficies que son punzantes, donde sus cuerpos frágiles pueden resultar heridos 57.
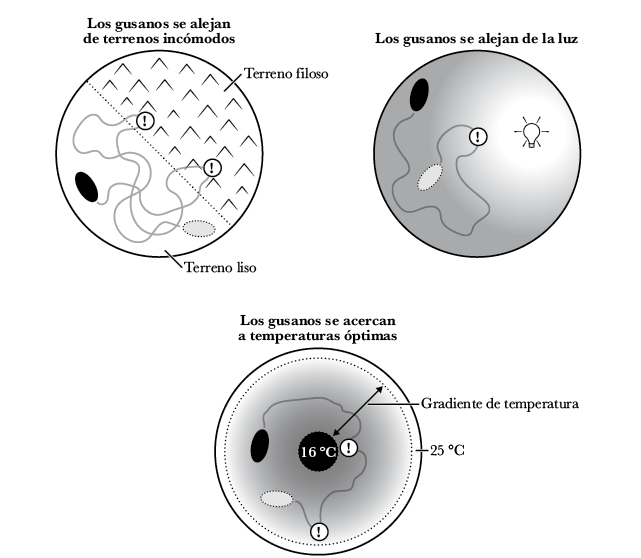
Figura 2.7: Ejemplos de decisiones de direccionalidad hechas por bilaterales simples como nematodos y gusanos planos
La estrategia de moverse con direccionalidad no era nueva. Los organismos unicelulares como las bacterias se mueven en sus entornos de una manera similar. Cuando un receptor de proteínas en la superficie de una bacteria detecta un estímulo como la luz, puede desencadenar un proceso químico dentro de la célula que cambie el movimiento de los propulsores de proteínas y, de ese modo, causará que cambie su dirección. Esta es la forma en que organismos unicelulares como las bacterias pueden acercarse a las fuentes de alimento o alejarse de químicos peligrosos. Sin embargo, ese mecanismo solo funciona en la escala de células individuales, donde los simples propulsores de proteínas pueden reorientar con éxito la forma de vida entera. Direccionar un organismo que contiene millones de células requiere una configuración completamente nueva, una en la que un estímulo active circuitos de neuronas y las neuronas activen células musculares, lo que al final provoca movimientos de giro específicos. Por lo tanto, el avance que trajo consigo el primer cerebro no era la direccionalidad per se, sino la direccionalidad en la escala de organismos pluricelulares.
El primer robot
En las décadas de 1980 y 1990 surgió una escisión en la comunidad de la inteligencia artificial. Por un lado, estaban aquellos situados en el campo de la IA simbólica, que se centraban en descomponer la inteligencia humana en sus partes constituyentes para intentar dotar a los sistemas de IA de nuestras habilidades más preciadas: el razonamiento, el lenguaje, la capacidad de resolución de problemas y la lógica. En oposición, se encontraban aquellos situados en el campo de la IA conductual, dirigidos por el experto en robótica Rodney Brooks del Instituto Tecnológico de Massachusetts, que creía que el enfoque simbólico estaba condenado al fracaso porque «nunca comprenderemos cómo descomponer el nivel de inteligencia humana hasta que no hayamos practicado mucho con niveles de inteligencia más simples».
El argumento de Brooks estaba basado en parte en la evolución: fueron necesarios mil millones de años solo para que la vida pudiera sentir y responder a su entorno; fueron necesarios otros quinientos mil millones de años de experimentación para que los cerebros se volvieran hábiles con las capacidades motoras y los movimientos. Y solo tras todo este arduo trabajo aparecieron el lenguaje y la lógica. Para él, en comparación con cuánto tiempo llevó desarrollar los sentidos y el movimiento, la lógica y el lenguaje aparecieron en un abrir y cerrar de ojos. Por lo tanto, concluyó que:
El lenguaje… y el razonamiento son muy simples una vez que la esencia de ser y la reacción se encuentran disponibles. Esa esencia es la capacidad de moverse en un entorno dinámico, percibir el entorno hasta un grado suficiente como para lograr el mantenimiento necesario de la vida y su reproducción. Esta parte de la inteligencia es donde la evolución ha concentrado su tiempo; es mucho más difícil.
Para Brooks, si bien los humanos «nos brindaron una prueba existente [de inteligencia a nivel humano], debemos ser cautelosos con las lecciones que se pueden extraer de ella» 58. Para ilustrar esto, ofreció una metáfora:
Supongamos que es la década de 1890. El vuelo artificial es el tema preferido en los círculos científicos, de ingeniería y de capital de riesgo. Un grupo de investigadores [de vuelos artificiales] logran transportarse de forma milagrosa con una máquina del tiempo a la década de 1990 durante algunas horas. Pasan todo el tiempo en la cabina de pasajeros de un Boeing 747 comercial durante un vuelo de duración media. Ya de regreso a la década de 1890, se sienten animados porque saben que [el vuelo artificial] es posible a gran escala. De inmediato comienzan a trabajar para duplicar lo que han visto. Avanzan con éxito en el diseño de asientos reclinables, ventanas de doble panel, y saben que, si tan solo pueden descifrar esos «plásticos extraños», tendrán el Santo Grial en sus manos 59.
Al intentar saltarse los aviones simples y construir directamente un 747, se arriesgaron a no comprender por completo los principios de cómo funcionan los aviones (los asientos reclinables, los cristales de las ventanas y los plásticos no eran los elementos en los que debían fijarse). Brooks creía que el ejercicio de aplicar la ingeniería inversa para comprender el cerebro humano era víctima de ese mismo problema. Un mejor enfoque era «construir las capacidades de los sistemas de inteligencia de forma que se fueran incrementando y lograr sistemas completos en cada paso». En otras palabras, comenzar como lo hizo la evolución: partir de cerebros simples y agregar complejidad desde ese punto.
Muchos no están de acuerdo con el enfoque de Brooks, pero ya sea que estés de acuerdo con él o no, fue el primero en construir un robot doméstico comercialmente exitoso; Brooks dio el primer pequeño paso hacia Robotina. Y en ese primer paso en la evolución de los robots comerciales podemos observar un paralelismo con la evolución del cerebro. Brooks, también, comenzó con la direccionalidad.
En 1990, cofundó una empresa de robótica llamada iRobot y, en 2002, introdujo a «Roomba», la primera aspiradora robot. La Roomba era un robot que recorría de manera autónoma tu casa aspirando el suelo. Fue un éxito inmediato; hoy en día todavía se producen, y la compañía ha vendido más de cuarenta millones de unidades 60.
La primera Roomba y los primeros bilaterales comparten un número sorprendente de características. Ambos tenían sensores extremadamente simples; la primera Roomba podía detectar tan solo algunas pocas cosas, como cuando golpeaba una pared o se encontraba en las cercanías de su base de carga. Ambos poseían cerebros simples; ninguno utilizaba la escasa entrada sensorial que recibía para dibujar un mapa de su entorno y reconocer objetos 61. Ambos eran bilateralmente simétricos. Las ruedas de la Roomba le permitían moverse solo hacia adelante y hacia atrás. Para cambiar de dirección, debía girar en su lugar y luego continuar con su avance hacia adelante.

Figura 2.8: Roomba. Una aspiradora robot de limpieza que se trasladaba de manera similar a los primeros bilaterales 62
La Roomba podía limpiar las rendijas y escondrijos de tu suelo solo con moverse de manera aleatoria, alejándose de objetos cuando se topaba contra ellos y acercándose a su base de carga cuando se le terminaba la batería. En cuanto chocaba contra una pared, daba un giro aleatorio y volvía a intentar moverse hacia adelante. Cuando tenía batería baja, buscaba una señal de su base de carga y, al detectarla simplemente giraba hacia donde la señal era más intensa y acababa regresando a su base de carga.
Las estrategias de desplazamiento de la Roomba y de los primeros bilaterales no eran idénticas, pero quizás no sea coincidencia que el primer robot doméstico exitoso se valiera de una inteligencia no tan diferente de la de los primeros cerebros. Ambos utilizaban estrategias que les permitían moverse en un mundo complejo sin comprender o modelar de verdad ese mundo.
Mientras que otros permanecían atrapados en el laboratorio trabajando en robots millonarios que tenían ojos, tacto y cerebros que intentaban calcular cuestiones complicadas como mapas y movimientos, Brooks construyó el robot más simple posible, uno que a duras penas tenía sensores y que no calculaba nada en absoluto. No obstante, el mercado, como la evolución, premia tres cosas por sobre todo lo demás: cosas que son baratas, cosas que funcionan y cosas que tienen la simpleza suficiente para ser descubiertas en primer lugar.
Si bien la direccionalidad quizás no inspire la misma admiración que otros logros intelectuales, sin dudas era energéticamente barata, funcionaba, y tenía la simpleza suficiente para que los experimentos evolutivos se toparan con ella. Desde ese lugar partieron los cerebros.
La valencia y el interior del cerebro de un nematodo
En la cabeza de un nematodo existen neuronas sensoriales, algunas de las cuales responden a la luz, otras al tacto, y otras a químicos específicos. Para que el control de dirección funcione, los primeros bilaterales debían considerar cada olor, cada roce u otro estímulo que detectaran y tomar una decisión: ¿me acerco, lo evito o lo ignoro?
El avance del control de dirección requirió que los bilaterales categorizaran el mundo en cosas a las que acercarse («cosas buenas») y cosas de las que alejarse («cosas malas»). Incluso la Roomba hace lo mismo: los obstáculos son malos; la base de carga cuando la batería es baja es buena. Antes, los animales radialmente simétricos no se movían, por lo que nunca tuvieron que categorizar al mundo de esta manera.
Cuando los animales categorizan los estímulos en buenos o malos, los psicólogos y neurocientíficos afirman que están dotando a los estímulos con valencia. «Valencia» es la bondad o maldad de un estímulo. No se trata de un juicio moral; es algo mucho más primitivo: se trata de si un animal responderá a un estímulo acercándose o alejándose de él. La valencia de un estímulo no es, por supuesto, objetiva; un químico, imagen o temperatura, en sí mismos, no poseen bondad o maldad. En cambio, la valencia de un estímulo es subjetiva y solo se define según la evaluación del cerebro sobre su bondad o su maldad.
¿Cómo establece un nematodo la valencia de algo que percibe? No realiza primero una observación cuidadosa y luego decide su valencia. En cambio, las neuronas sensoriales de su cabeza señalan directamente la valencia del estímulo. Un grupo de neuronas sensoriales son, en efecto, neuronas de valencia positiva; se activan directamente con cosas que los nematodos consideran buenas (como olores a comida). Otro grupo de neuronas sensoriales son, en efecto, neuronas de valencia negativa; se activan directamente con cosas que los nematodos consideran negativas (como altas temperaturas, olor a depredadores, luz intensa).
En los nematodos, las neuronas sensoriales no señalan características objetivas del mundo que los rodea; codifican «votos de dirección» sobre cuánto desea un nematodo acercarse o alejarse de algo. En bilaterales más complejos, como los seres humanos, no todo el mecanismo sensorial funciona de esta manera. Las neuronas de tus ojos detectan características de las imágenes; la valencia de la imagen se calcula en otro lugar. No obstante, al parecer, los primeros cerebros comenzaron con neuronas sensoriales que no se preocupaban por medir las características objetivas del mundo y, en cambio, interpretaban toda la percepción mediante el simple lente binario de la valencia.
La figura 2.9 enseña un diagrama simplificado de cómo funciona la direccionalidad en los nematodos. Las neuronas de valencia desencadenan diferentes decisiones de giro al conectarse con diferentes neuronas descendentes 63.
Consideremos cómo utiliza un nematodo este circuito para encontrar alimento. Los nematodos poseen neuronas de valencia positiva que desencadenan el movimiento hacia adelante cuando se intensifica la concentración del olor del alimento. Como ya vimos con las neuronas sensoriales en la red nerviosa de los primeros animales, estas neuronas se adaptan rápidamente a los niveles base de los olores. Esto les permite a estas neuronas de valencia señalar los cambios en un rango amplio de concentración de olores. Estas neuronas generarán un número similar de picos tanto si la concentración del olor sube de dos a cuatro partes o de cien a doscientas partes. Esto hace que las neuronas de valencia puedan seguir empujando al nematodo en la dirección correcta. Es la señal de «¡sí, sigue avanzando!» del primer olfateo de un alimento lejano hasta la fuente del alimento en sí mismo.

Figura 2.9: Esquema simplificado de la conexión del primer cerebro
Este uso de la adaptación es un ejemplo de cómo las innovaciones evolutivas permiten innovaciones futuras. La capacidad de acercarse a la comida de los primeros bilaterales fue posible solo porque la adaptación ya había evolucionado en los primeros animales radialmente simétricos. Sin la adaptación, las neuronas de valencia serían muy sensibles (y se activarían de forma continua cuando los olores estuvieran demasiado cerca) o no tendrían la sensibilidad suficiente (serían incapaces de detectar olores lejanos).
En este punto, podrían aparecer nuevos comportamientos de dirección con solo modificar las condiciones por las cuales se excitan las diferentes neuronas de valencia. Por ejemplo, consideremos cómo se acercan los nematodos hacia temperaturas óptimas.
El movimiento con respecto a la temperatura requiere una astucia adicional con respecto al simple movimiento hacia los olores: la concentración menor de un alimento siempre es mala, pero el decrecimiento de la temperatura de un entorno solo es malo si el nematodo ya siente demasiado frío. Si un nematodo siente calor, entonces la disminución de la temperatura es buena. Un baño caliente resulta insoportable en un verano abrasador; sin embargo, se convierte en una bendición en un invierno helado. ¿Cómo lograron los primeros cerebros gestionar las fluctuaciones de temperatura de manera diferente dependiendo del contexto?
Los nematodos cuentan con una neurona de valencia negativa que provoca el giro cuando las temperaturas aumentan, pero solo si la temperatura ya se encuentra sobre un umbral determinado; es una neurona de «¡demasiado caliente!». Los nematodos también tienen una neurona de «¡demasiado frío!»; activa el giro cuando las temperaturas bajan, pero solo cuando ya se encuentran por debajo de cierto umbral. Juntas, estas dos neuronas de valencia negativa permiten que los nematodos se alejen rápidamente del calor cuando sienten demasiado calor y se alejen del frío cuando sienten demasiado frío 64. En las profundidades del cerebro humano hay una estructura antigua llamada «hipotálamo», hogar de las neuronas sensibles a la temperatura, que funciona de la misma manera.
El problema de realizar concesiones
La direccionalidad en presencia de múltiples estímulos presentaba un problema: ¿qué sucede si diferentes células sensoriales optan por moverse en direcciones opuestas? ¿Qué sucede si un nematodo siente el aroma de algo delicioso y algo peligroso al mismo tiempo?
Los científicos han puesto a prueba a los nematodos justo en esa clase de situaciones. Reúne a un grupo de nematodos a un lado de una placa de Petri y coloca un alimento delicioso en el lado opuesto de la placa; luego coloca una peligrosa barrera de cobre (los nematodos odian el cobre) en el medio. Los nematodos se enfrentan a un problema: ¿están dispuestos a atravesar la barrera para obtener la comida? De manera sorprendente, la respuesta es —tal como cabría esperar de un animal con siquiera un mínimo de inteligencia— que depende. Depende de la concentración relativa del alimento en contraposición al aroma del cobre.
Si el nivel del cobre es bajo, la mayoría de los nematodos cruzan la barrera; si el nivel de cobre es intermedio, solo algunos deciden cruzar. Frente a un nivel alto de cobre, ningún nematodo está dispuesto a cruzar la barrera 65.
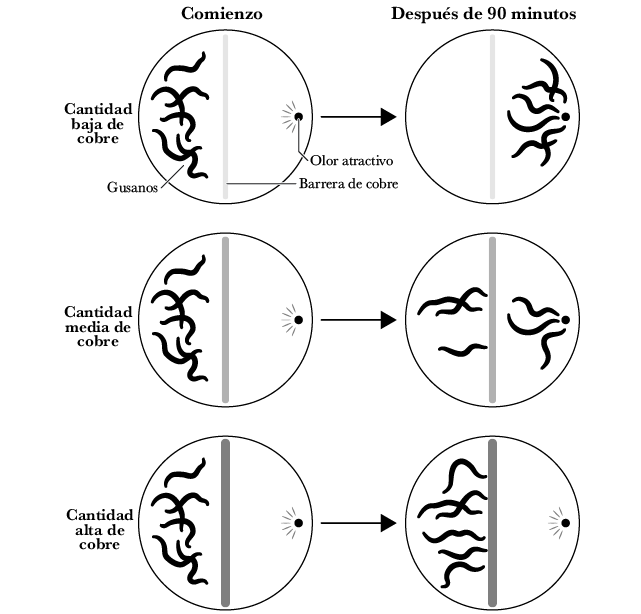
Figura 2.10
Esta capacidad de establecer concesiones en el proceso de toma de decisiones ha sido evaluada en diferentes especies de bilaterales de la familia de los gusanos y teniendo en cuenta diferentes modalidades sensoriales 66. Los resultados indican de manera constante que incluso los cerebros más simples, aquellos con menos de mil neuronas, pueden realizar concesiones.
Es posible que este requerimiento de integrar la entrada de datos a lo largo de las modalidades sensoriales haya sido una razón por la cual la direccionalidad necesitó de un cerebro y no podría haber sido implementada en una red distribuida de reflejos como la de los pólipos coralinos. Todas estas entradas sensoriales que votaban para moverse en diferentes direcciones debían ser integradas en conjunto en un único lugar para tomar una única decisión; solo puedes ir en una sola dirección a la vez. El primer cerebro era un centro de megaintegración, un gran circuito neuronal en donde se seleccionaban las direcciones de movimiento.
Puedes intuir cómo funciona esto a partir de la figura 2.9, que enseña una versión simplificada del circuito de dirección de los nematodos. Las neuronas de valencia positiva se conectan a una neurona que desencadena el movimiento hacia adelante (lo que se podría denominar «neurona de avance»). Las neuronas de valencia negativa se conectan a una neurona que desencadena el giro (lo que se podría denominar «neurona de giro»). La neurona de avance acumula votos de «¡sigue avanzando!» y la neurona de giro acumula votos de «¡cambia de dirección!». Las neuronas de avance y giro se inhiben la una a la otra, lo que permite que esta red incorpore concesiones y tome una sola decisión; la neurona que acumule más votos gana y determina si el animal cruzará la barrera de cobre 67.
Este es otro ejemplo de cómo las innovaciones pasadas permitieron innovaciones futuras. Al igual que un bilateral no puede avanzar y girar al mismo tiempo, un pólipo coralino no puede abrir y cerrar la boca en un mismo movimiento. Las neuronas inhibitorias evolucionaron en los primeros animales coralinos para permitir que estos reflejos mutuamente exclusivos compitieran entre sí para que solo un reflejo pudiera ser seleccionado a la vez; este mismo mecanismo fue reutilizado por los primeros bilaterales para permitirles realizar concesiones en sus decisiones de direccionalidad. En lugar de decidir si abrir o cerrar la boca, los bilaterales utilizaron a las neuronas inhibitorias para decidir si avanzar o girar.
¿Tienes hambre?
La valencia de algo depende del estado interno del animal. La decisión de un nematodo de cruzar una barrera de cobre para obtener alimento no depende solo del nivel relativo de intensidad del olor a comida y cobre, sino también de lo hambriento que se encuentre el nematodo 68. No cruzará la barrera para obtener comida si está satisfecho, pero lo hará si tiene hambre. Además, los nematodos pueden trastocar por completo sus preferencias según el hambre que tengan 69. Si un nematodo está bien alimentado, se alejará del dióxido de carbono; si tiene hambre, se acercará a él. ¿Por qué? El dióxido de carbono es un químico que liberan tanto la comida como los depredadores, de modo que, cuando un nematodo está satisfecho, dirigirse hacia el dióxido de carbono para obtener alimento no compensa el riesgo de enfrentarse a un depredador. No obstante, cuando tiene hambre, la posibilidad de que el dióxido de carbono esté indicando la presencia de comida y no de depredadores hace que valga la pena arriesgarse.
La capacidad del cerebro de cambiar a toda velocidad la valencia de un estímulo dependiendo de los estados internos es ubicua. Comparemos el éxtasis salival del primer bocado de tu cena favorita después de un largo día en el que te saltaste comidas con la sensación de náuseas e hinchazón tras el último bocado y de haber engullido hasta el cansancio. En cuestión de minutos, tu comida favorita puede pasar de ser un regalo de Dios a la humanidad a algo que no quieres ni siquiera oler.
Los mecanismos por lo que esto sucede son relativamente simples y se presentan en todos los bilaterales. Las células animales liberan químicos específicos —«señales de saciedad»; por ejemplo, la insulina— como respuesta a haber alcanzado una cantidad saludable de energía 70. Y las células animales liberan un conjunto diferente de químicos —«señales de hambre»— como respuesta a una cantidad insuficiente de energía. Ambas señales se esparcen por el cuerpo del animal y proveen una señal global persistente que indica su nivel de hambre. Las neuronas sensoriales de los nematodos cuentan con receptores que detectan la presencia de esas señales y cambian sus respuestas en consecuencia. Las neuronas de valencia positiva con respecto al olor de la comida en los C. elegans se vuelven más receptivas en presencia de señales de hambre y menos receptivas en presencia de señales de saciedad.
Los estados internos también se encuentran presentes en la Roomba, ya que ignorará la señal de su base de carga cuando su batería esté completa. En este caso, se podría afirmar que la señal de la base de carga es de valencia neutral. Cuando su estado interno cambia a un estado de batería baja, la señal de la base de carga cambia y adquiere valencia positiva: la Roomba ya no ignorará la señal de su base de carga y se acercará a ella para cargar su batería.
La direccionalidad requiere al menos cuatro aspectos: un plan corporal bilateral para girar, neuronas de valencia para detectar y categorizar estímulos en buenos o malos, un cerebro para integrar las entradas de información y tomar una sola decisión de dirección, y la capacidad de modular la valencia teniendo en cuenta los estados internos. Pero, aun así, la evolución continuó experimentando. Existe otra estrategia que surgió en los primeros cerebros bilaterales, una estrategia que fortaleció aún más la eficacia del control direccional. Esta estrategia fue el origen de lo que ahora denominamos «emoción».