Principios de la multiplicación por semillas
Reproducción sexual
La base de la reproducción sexual (gámica) de las plantas es la meiosis celular, que es el estadio esencial de la formación de las células reproductoras. En el curso de la meiosis, el patrimonio genético de los padres queda reducido a la mitad por la separación de los genes homólogos de cada una de las parejas de cromosomas; así, una célula de polen (masculina) o un oocito (femenina) contienen sólo la mitad del patrimonio genético de su progenitor.
Al término de este proceso se forma un huevo maduro, que podrá ser fecundado y dar una semilla. La fecundación se produce gracias a la polinización, por obra del viento, de los animales (insectos, pájaros...) o del hombre. Esta se realiza casi siempre entre plantas de la misma especie, más raramente entre plantas del mismo género y casi nunca entre plantas de géneros diferentes.
Mediante la fecundación, los genes de las plantas, contenidos en los gametos, forman una nueva combinación. Un carácter únicamente se encontrará con toda seguridad en los descendientes si los padres son homocigóticos en un carácter determinado; si no es así, que es lo que pasa la mayor parte de las veces, es decir, si los padres son heterocigóticos o genéticamente diferentes en un carácter determinado, su descendencia podrá manifestar este carácter de forma distinta según la manera en que se hayan combinado los genes y según sea el carácter: dominante o recesivo.

Clavel interespecífico. (©Biosphoto/Harold Verspieren/Digitalice)

Flor de un cultivar de nenúfar. (©Biosphoto/Pascal André)
Los individuos provenientes de la unión de plantas de la misma especie pero que tienen caracteres diferentes, y los que proceden de especies distintas del mismo género se denominan híbridos.
Los híbridos con padres de la misma especie (híbridos interespecíficos) generalmente son fértiles, mientras que los de especies diferentes del mismo género (híbridos intergenéricos) suelen ser estériles. Por consiguiente, la reproducción por semillas, que constituye la reproducción sexual por excelencia, permite que los padres puedan transmitir a su descendencia infinitas variables del genotipo y fomenta así la diversidad.
En el plano estético, las plantas hijas pueden parecerse de una manera más o menos aproximada a uno u otro progenitor.
Cuando, gracias a esta variabilidad, las plantas nuevas presentan unas características totalmente diferentes de los padres, se obtiene una nueva variedad. Las variedades nuevas pueden aparecer en la naturaleza o puede ser creadas por el ser humano por hibridación, en cuyo caso reciben la denominación de cultivar (del inglés cultivated variety).
Pero si quisiera diseminarse esta nueva variedad (este cultivar), no podría recurrirse a la reproducción sexual y utilizar las semillas (a excepción de los híbridos de primera generación provenientes de líneas puras: véase «Creación de nuevos cultivar), sino que debería emplearse en este caso la multiplicación vegetativa (esquejes, injertos, acodos, amugronamiento, etc.).
El origen de este proceso no es la meiosis, sino que se trata de la mitosis.
La mitosis es el modelo usual de división de célula viva que asegura el mantenimiento del mismo número de cromosomas.
En la mitosis, los cromosomas se reproducen dividiéndose longitudinalmente, pero no se combinan entre sí par producir algo nuevo. Por lo tanto, el patrimonio genético de la planta progenitora, con todas sus características estéticas y morfológicas, se transmite en su totalidad a las células hijas sin que haya modificaciones en la descendencia.
Del fruto a la semilla
En las angiospermas (plantas que tienen un ovario floral), cuando la flor ha sido fecundada, el óvulo se modifica, aumenta de tamaño y da la semilla. A partir de ese momento, el ovario que la contiene se comienza a transformar, de modo diferente según las especies, en fruto.
El conjunto de la semilla y la evolución del ovario recibe el nombre de fruto verdadero; por ejemplo, la cereza es un fruto verdadero. Pero en ciertos casos los tejidos del receptáculo también se transforman, se hacen carnosos y dan lugar a un falso fruto; la manzana es un ejemplo de ello. A veces, lo que se presenta como un fruto en realidad es una infrutescencia, formada por varios frutos agrupados; es el caso de la frambuesa.
En las gimnospermas (plantas cuya flor no posee ovario), la situación es totalmente diferente: los «frutos» de las coníferas (las bayas, las piñas o, para emplear el término exacto, los conos, etc.) están hechos de escamas, que se han desarrollado y lignificado después de que las flores hayan sido fecundadas con la función de proteger los óvulos.
En las angiospermas se distinguen, por un lado, los frutos carnosos y, por otro, los frutos secos.
Frutos carnosos
En los frutos carnosos se diferencian tres partes: el epicarpio, es decir, la parte externa (la piel); el mesocarpio, que es la zona intermedia, carnosa y jugosa, y el endocarpio, en el interior, que contiene la semilla.
Los frutos carnosos propiamente dichos son las drupas, como las cerezas o los melocotones (que tienen una sola semilla), y las bayas, como la uva (con varias semillas).
Los falsos frutos son los piridiones (manzanas y peras), las hespérides (cítricos) y los compuestos (moras y frambuesas).

Piña. (©Biosphoto/Fève Frédéric)

Frambuesa. (©Biosphoto/Whitworth Jo/GAP)
Frutos secos
Los frutos secos se dividen, por un lado en frutos monocárpicos, los cuales contienen una sola semilla, y, por tro, frutos policárpicos, con varias semillas.
A su vez, estos dos tipos pueden subdividirse en frutos indehiscentes (que son aquellos se mantienen cerrados) y frutos dehiscentes (que son los que se abren al madurar).
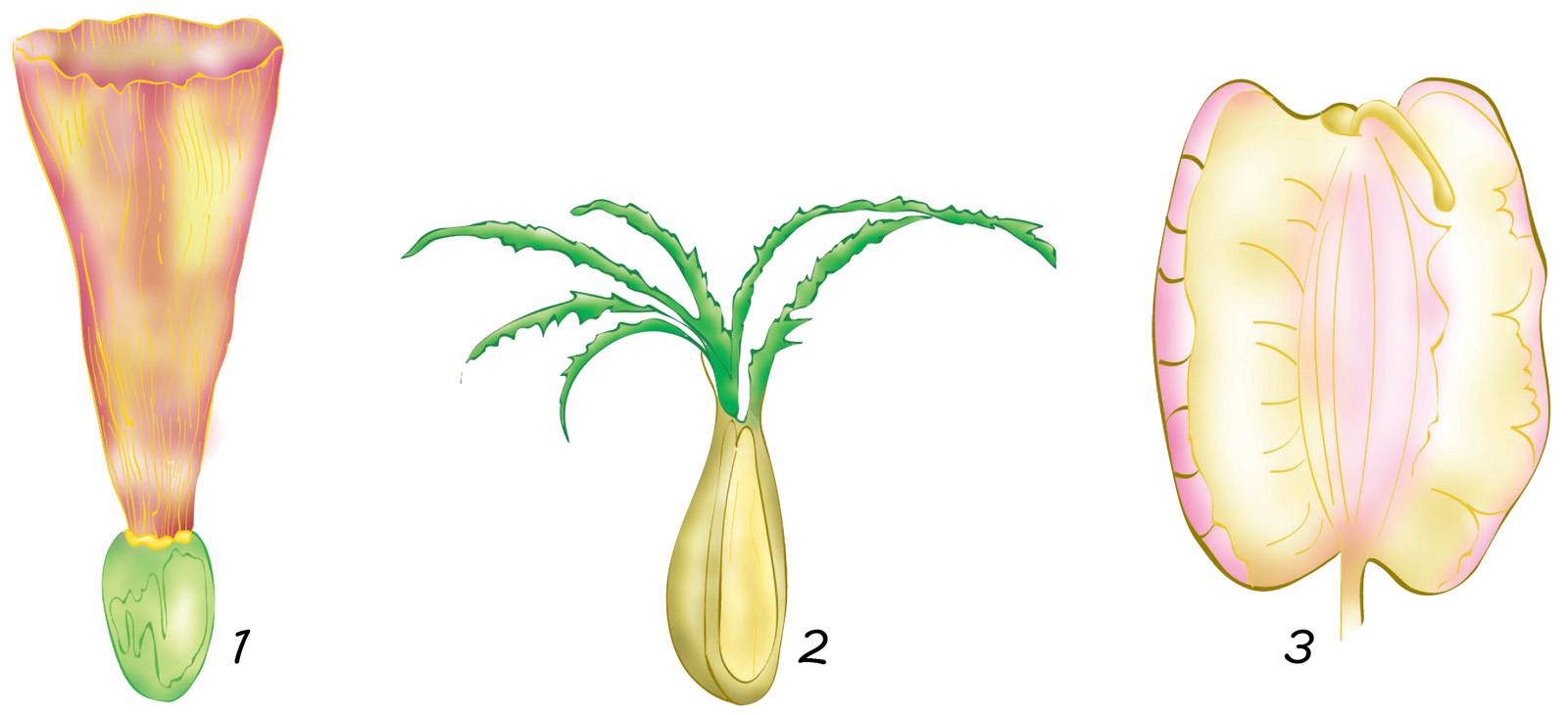
Entre los indehiscentes están los aquenios (de la familia Asteráceas, antes denominada Compuestas), las sámaras (olmos, arces y fresnos), las cariopsis y las nueces (robles, nogales y castaños).
Los frutos dehiscentes son:
— las legumbres (Leguminosas);
— las cápsulas (amapolas, Impatiens);
— los folículos (eléboros, ranúnculos;
— las silicuas (de la familia Brasicáceas, antes llamada Crucíferas).
En las cápsulas, las semillas se liberan de varias maneras:
— Impatiens, en este sentido, es muy característica debido a que proyecta sus granos a distancias considerables;
— en otras especies, como las amapolas, las semillas salen por unos orificios.
Los folículos suelen contener una sola semilla, mientras que en el caso de las legumbres es habitual que cuenten con varias.
Estos frutos pueden tener formas muy diferentes: rectas, en espiral, con una cavidad única o con secciones que separan las semillas.
1. Aquenio de alcachofa; 2. Aquenio de valeriana; 3. Fruto alado de angélica
Semillas no comestibles
Las semillas de algunas plantas son venenosas o, en cualquier caso, resultan tóxicas cuando se ingieren. Entre las plantas ornamentales se encuentran la glicinia, el codeso, Delphinium y el tejo; del resto de las plantas, el ricino es particularmente tóxico.
Las semillas de ciertos frutos, ingeridas en gran cantidad, también pueden tener efectos mortales: es el caso de las semillas de la manzana, del melocotón, de la ciruela y de la cereza. En todos los casos, se aconseja no dejar las semillas al alcance de los niños.
Formas, tamaños y colores de las semillas
Las semillas son de formas y tamaños muy variables. Hay semillas redondas, ovoides, oblongas, reniformes, en forma de disco, etc.
El tamaño de las semillas no siempre es directamente proporcional a las dimensiones de la planta que después originarán.
Por ejemplo, las semillas de una de las especies de árbol más grandes del mundo, Sequoia sempervirens, son pequeñas, estrechas y están en unas piñas (estróbilos) de apenas 2 cm de largo, formadas por 15 o 20 escamas, que contienen, cada una de ellas, de 3 a 7 óvulos. Un kilo de estas semillas equivale a unas 230 000 semillas; en cambio, un kilo de Pinus pinea contiene unas 1 250 semillas (piñones).
Algunas semillas tienen colores especialmente bonitos; por ejemplo, la semilla de color naranja vivo de Cycas revoluta, la semilla vellosa de tonalidad azul cielo de Ravenala madagascarensis o la semilla negra y brillante de Paeonia delavayi. Las vainas también pueden presentar una amplia gama de colores llamativos, como por ejemplo las bayas rojo vivo de Erythina crista galli.

Primer plano de una flor de adormidera. (©Biosphoto/Jariwala Hemant/GPL)

1. Legumbre de loto; 2. Cápsula de violeta
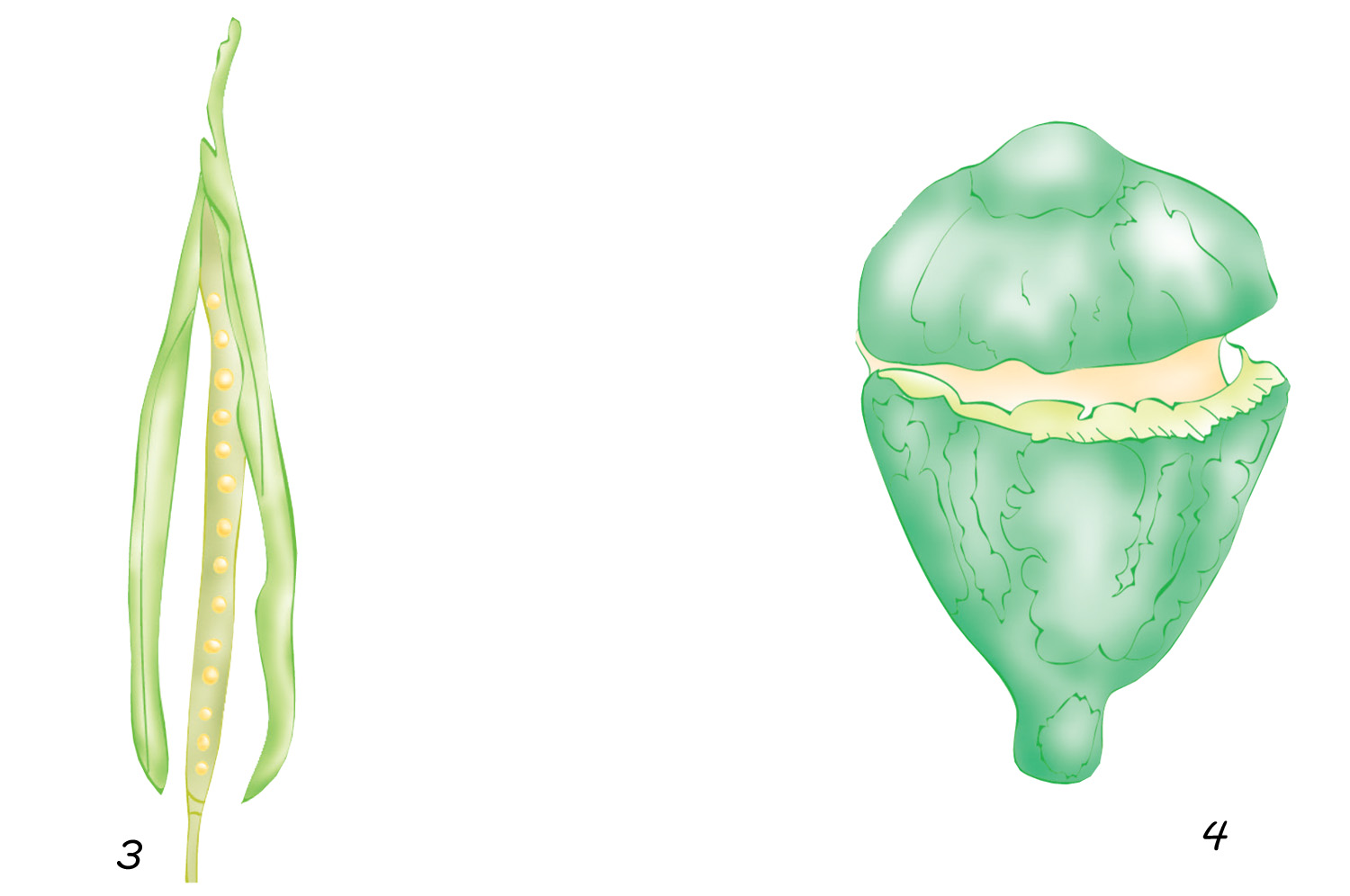
3. Silicua de brasicáceas; 4. Cápsula de eucalipto

Semillas de girasol rojo. (©Biosphoto/Giraud Philippe)

Semillas de girasol. (©Biosphoto/Giraud Philippe)
Sustancias de reserva
En la mayor parte de los casos, las semillas tienen una reserva de sustancias nutritivas que sirve para que el embrión se mantenga vivo durante la fase de germinación. Sin embargo, hay semillas que no poseen ninguna reserva: son de un tamaño muy pequeño, con un tegumento muy delicado, y se producen en grandes cantidades. Son ejemplos de este tipo las semillas de las orquídeas, de las bromeliáceas o del brezo.
En las dicotiledóneas, las sustancias de reserva se encuentran en las hojas del mismo nombre, que son las primeras que se desarrollan después de la germinación. Estas hojas son, por forma y tamaño, muy diferentes al resto. En las monocotiledóneas, el cotiledón tiene la función de absorber las sustancias nutritivas. El embrión de estas plantas es muy pequeño y hundido, y las sustancias nutritivas están en la semilla.
El cocotero de las Seychelles
La semilla más grande del mundo es la que produce una palmera muy parecida al cocotero, Lodoicea maldivica.
Sus gigantescas semillas, grandes como dos cocos, aparecían a menudo varadas en la arena de playas bañadas por aguas del Índico pero, al tratarse de semillas no vitales, no podía comprobarse de qué árboles procedían; incluso se creía que el árbol que las daba crecía bajo las aguas del océano. De ahí que recibiera el nombre de cocotero de mar.
Hasta finales del siglo XVIII no se supo que estas semillas provenían de una palmera que crece en una región muy delimitada de las islas Seychelles.
Se trata de un árbol que puede llegar a alcanzar los 30 m de altura, poniendo el tronco recto y liso. Sus hojas tienen forma de abanico cerrado y pueden medir hasta 6 m.
Es una especie dioica, es decir, tiene flores masculinas y flores femeninas en plantas separadas.
Los individuos hembra son más grandes y producen cocos que pesan hasta 20 kg, que tardan casi siete años en madurar.
En cambio, los ejemplares masculinos suelen ser más pequeños y es habitual que den inflorescencias en forma de espiga, de color chocolate.
Estas semillas tienen la pulpa llena: esto explica por qué, a diferencia de los cocos normales (Cocos nucifera), que son huecos por dentro, los cocos de Lodoicea no flotan y acaban muriendo en las aguas saladas.
Por esta razón, mientras el cocotero ha aumentado su área de difusión gracias a que sus semillas flotan y ello les permite recorrer largas distancias, Lodoicea ha permanecido en una zona muy limitada.

(©Biosphoto/Martin Gilles)
Estructura de la semilla
Las semillas de formas y tamaños diferentes tienen, sin embargo, en común que están divididas en tres partes distintas: embrión, endosperma y tejidos protectores.
Embrión
Es la parte que dará lugar a una nueva planta. El embrión proviene de la fusión de los gametos masculinos y los gametos femeninos. Durante la formación de la semilla, el embrión crece hasta un cierto estadio, cuando alcanza la fase «durmiente».
En muchas especies, esta etapa puede prolongarse mucho tiempo: se ha descubierto que ciertas semillas estaban vivas incluso después de miles de años de haberse formado.
En un embrión resulta fácil apreciar las diferentes partes que originarán la nueva planta: la radícula, que generará todo el sistema radical; la plúmula, que formará el eje del tronco o del tallo, y las hojas primordiales, llamadas cotiledones.
Las gimnospermas poseen un número muy variable de hojas cotiledóneas, mientras que las angiospermas sólo tienen una o dos. Se dan las monocotiledóneas (las más extendidas son las de la familia Gramináceas, que son las hierbas que se encuentran comúnmente en los prados) y las dicotiledóneas (que forman la mayor parte de las latifolias).
La parte situada entre la radícula y los cotiledones se llama hipocótilo; en cambio, después de la germinación, la zona que se encuentra entre los cotiledones y las primeras hojas verdaderas recibe el nombre de epicótilo. En los brotes jóvenes, los cotiledones permiten la fotosíntesis y constituyen una importante reserva de alimento. Una vez que han cumplido su función, se marchitan y dejan su lugar a hojas verdaderas.
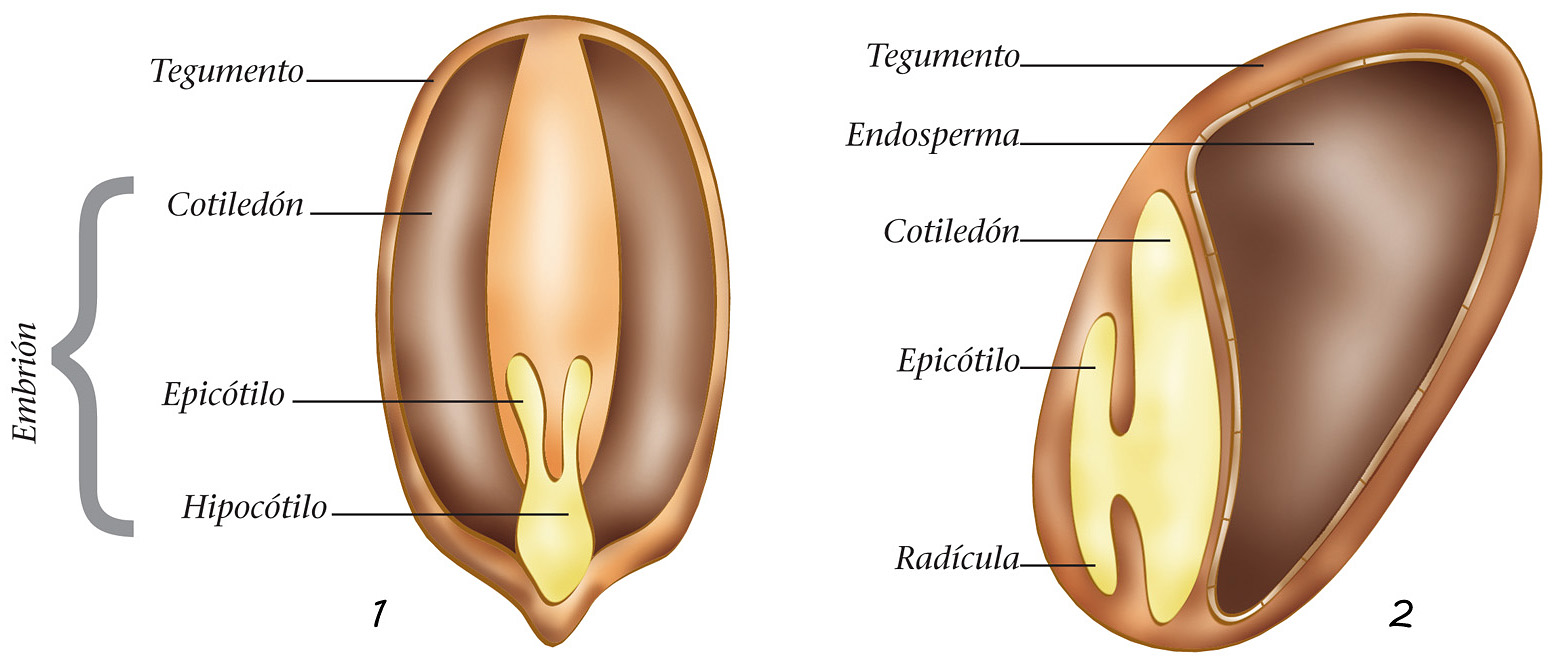
1. Semilla de planta dicotiledónea; 2. Semilla de monocotiledónea
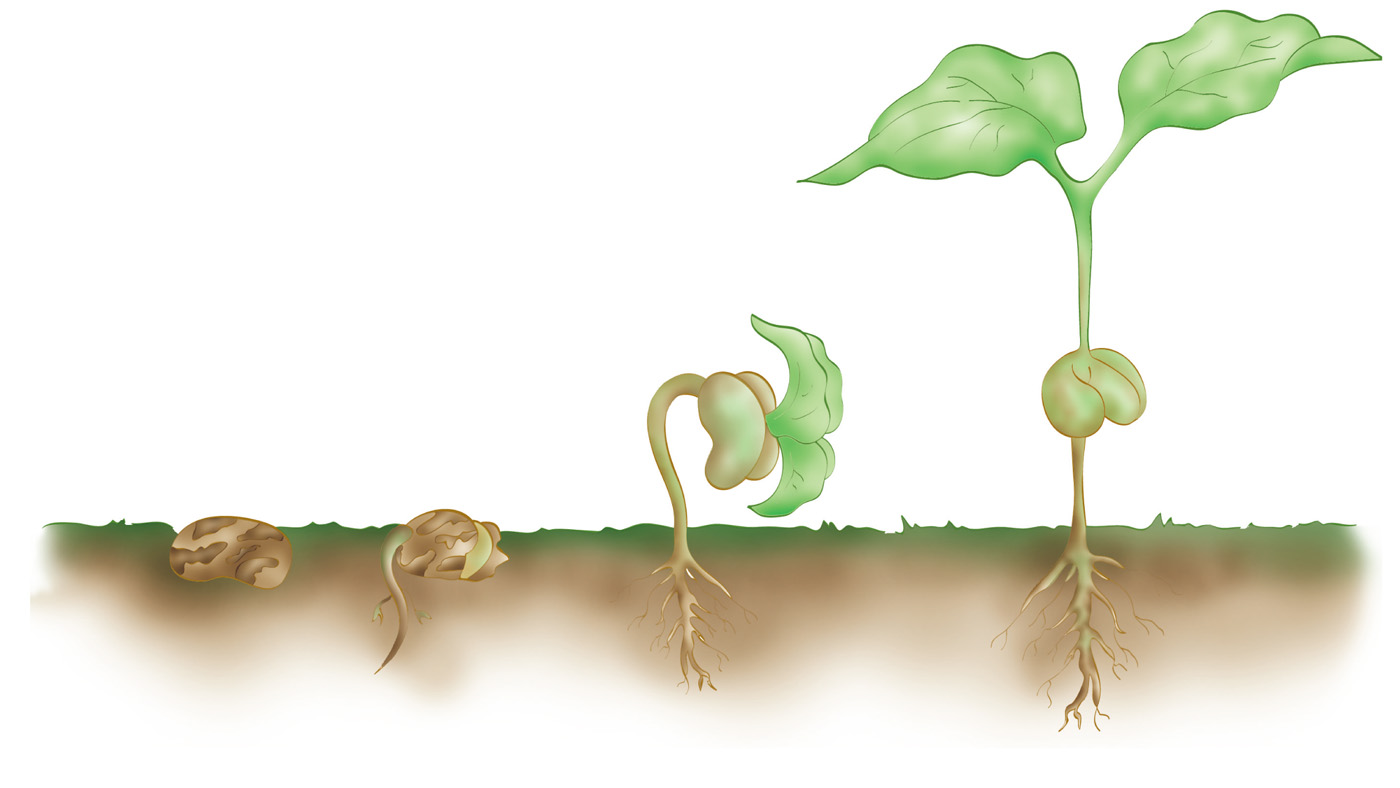
En una semilla de judía en fase de germinación, los cotiledones se aprecian claramente
Endosperma
Su función principal es convertirse en una reserva de alimento.
Contiene las sustancias que permiten la germinación de la semilla y el desarrollo de las plántulas (plantas muy jóvenes) mientras la fotosíntesis y la absorción de las sustancias nutritivas que aporta el suelo todavía están limitadas.
El origen del endosperma es diferente entre las gimnospermas, en las que proviene directamente del gametofito femenino, y las angiospermas, en las que sus tejidos nacen de la fusión de los gametos masculinos y femeninos.
La evolución de los vegetales hace que, en las semillas de las plantas que están consideradas evolucionadas, el endosperma sea muy pequeño o incluso no exista. En este caso, la función de reserva alimentaria está asegurada principalmente por los cotiledones.

Primer plano de un arilo de tejo común. (©Biosphoto/MG de Saint Venant)
Tejidos protectores
Envuelven el embrión para protegerlo. Provienen casi siempre de los tegumentos del óvulo y, a veces, de los del ovario.
Su principal función es impedir que el embrión se seque y también evitar que el agua penetre en la semilla antes de que esta se encuentre en condiciones aptas para germinar. Los tejidos protectores sufren varias transformaciones: pueden lignificarse,volverse carnosos o coger color y transformarse en comestibles para incitar a los animales a que tomen las semillas, a fin de favorecer su diseminación. En algunas plantas, como en el tejo (Taxus baccata), los tejidos tegumentarios pueden resistir los efectos de los jugos gástricos de los animales que los ingieren.
Diseminación
Las plantas que poseen semillas (espermatofitas) han adoptado, en función del medio en el que viven, «técnicas» particulares para distribuirlas y colonizar un territorio, según unas modalidades que a veces son realmente ingeniosas y espectaculares. Algunas se entregan a la acción del viento (anemocorias); otras, a la intervención de los animales (zoocorias), y otras propulsan directamente su descendencia a gran distancia. Por último, las hay que se sirven del agua como medio de distribución (hidrocorias).
En general, las modalidades de transporte corresponden a comportamientos diferentes según las semillas: las especies anemófilas normalmente no tienen periodo durmiente o, si lo tienen, este es ligero. Cuando, por el contrario, el transporte se realiza por medio de animales, el periodo durmiente es profundo y prolongado.
Hay que señalar que ciertas plantas no «quieren» que su progenitura se disperse: han encontrado un lugar donde se sienten bien y «creen» que su descendencia también estará a gusto allí. Sus flores, después de la fecundación y la desfloración, enrollan el pedúnculo y llevan al suelo sus frutos y sus semillas, que de este modo germinarán en la tierra donde la madre había encontrado un terreno idóneo.
Esto es lo que ocurre por ejemplo con los cacahuetes o, dentro del grupo de las plantas ornamentales, con los pamporcinos.
Tantas formas, tantas necesidades
La forma, el tamaño y las características de las semillas influyen, tal como se verá más adelante con mucho más detalle, en las diversas modalidades de diseminación.
Las semillas muy finas (orquídeas, begonia, tabaco ornamental, etc.) disponen de pocas sustancia nutritivas y corren el riesgo de perder rápidamente el poder germinativo; por ello deben sembrarse en la superficie y tiene que vigilarse mucho el riego durante las primeras fases de germinación, para no dispersarlas.
Las semillas con plumas (Gerbera, Gazania, Erigeron, etc.) tampoco tienen que sembrarse en profundidad, ya que la parte plumosa tiene que emerger del suelo.
En general, las semillas con un tegumento particularmente duro necesitan recibir un tratamiento (escarificación o estratificación) antes de poder germinar. Para evitar que estas semillas se sequen, debe prestarse una particular atención a las semillas oleaginosas (como las de la magnolia) durante la fase de conservación; si no pueden sembrarse de forma inmediata, deben conservarse en turba húmeda a baja temperatura.
Las semillas carnosas (como las del roble y del castaño) han de ponerse en agua durante 24 horas, sobre todo si las semillas que se pretenden cultivar son viejas.
En cuanto a las semillas con alas (semillas del arce y del fresno), deben desalarse para así repartirlas mejor durante la siembra y reducir el riesgo de marchitamiento.
La mayor parte de las semillas de plantas acuáticas deben conservarse en agua.
Una notable excepción la constituyen aquellas especies que no se sumergen después de la maduración (como sucede, por ejemplo, con el caso de la flor de loto), que se conservan en turba húmeda.
Anemocoria
Las semillas (o los frutos) que dispersa el viento deben ser muy ligeras o bien tener una forma que favorezca el «vuelo».
De este grupo pueden citarse las semillas del tabaco (incluido el tabaco ornamental) y las de las orquídeas, que se producen en abundancia, a veces incluso varios millones de semillas por pie.
Las semillas de Dendrobium pesan menos de una centésima de miligramo; son semillas minúsculas que no contienen sustancias nutritivas de reserva para la germinación.
Tienen formas curiosas y especiales: los vilanos, característicos de las Compuestas (por ejemplo, Taraxacum o diente de león), tienen forma de plumas o de pequeños paracaídas; las sámaras de los arces, los fresnos o los olmos recuerdan a las hélices de un helicóptero, o los frutos de los carpes, los tilos y los abedules.
Transportada por el viento, a veces incluso a unas alturas realmente vertiginosas, la descendencia de estas plantas puede aterrizar a varios kilómetros de distancia.

Diente de león en fruto y diseminación por el viento. (©Biosphoto/Hazan Muriel)
Barocoria
El caso contrario es el de ciertas plantas que producen semillas pesadas, que caen verticalmente al pie de la planta. Se dice que son barocorias, del griego baros, «peso», y khoreo, «desplazarse».
Las semillas más grandes del mundo son las de Lodoicea maldivica, que, como ya se ha visto, pueden llegar a pesar 20 kg.
En general, este tipo de diseminación tiene puntos débiles: la planta madre da sombra a las plantas jóvenes que están en su pie, y la acumulación de estas hace que la competencia entre ellas sea muy grande.
Algunas de las plantas con esta característica son los nogales, los robles y muchas plantas frutales.
Autocoria
Otras plantas han desarrollado un mecanismo que les permite proyectar sus semillas, una vez maduras, a distancias notables: es la autocoria o diseminación activa.
Un ejemplo de ello es Impatiens balsamina, también llamada brinco, que precisamente debe su nombre a esta característica. Al menor contacto, sus frutos en forma de porra se enrollan y expulsan con fuerza las semillas que contienen.
Zoocoria
Las plantas zoocorias se subdividen entre las que «viajan» en el interior del animal portador y las que se adhieren a él por el exterior. En el primer caso, los frutos de las plantas sirven de alimento a los animales; sus semillas, que poseen un tegumento resistente a los jugos gástricos del animal, se expulsan junto con los excrementos a una distancia y en unos lugares adonde difícilmente hubiesen podido llegar de otro modo.
Son zoocorias las plantas con bayas de vivos colores (acebos, tejos, Cotoneaster, Euonymus, etc.) que atraen a los pájaros en periodos en los que empieza a escasear la comida.
Un caso particular de zoocoria es la mirmecocoria, es decir, el transporte de semillas por medio de hormigas. Una de las plantas que utilizan a estos insectos para trasladar sus semillas es la celidonia o hierba verruguera (Chelidonium majus): sus semillas tienen unas crestas oleaginosas que gustan mucho a las hormigas.
Otra forma de zoocoria es aquella de las semillas que, dotadas de aguijones o ganchos, se adhieren al pelo de los animales. Algunas de estas plantas son el lampazo (Arctium lappa) y el amor de hortelano (Galium aparine).
El ser humano también puede contribuir, sin ser consciente de ello, al transporte de las semillas cuando las lleva adheridas a su ropa; es el caso de Circaea lutetiana o Bidens tripartita.
También se han registrado casos de semillas pegadas a paquetes transportados de un continente a otro, como ha ocurrido con Erigeron annuus y Solidago canadensis.

Caballo de la Camarga con frutos de lampazo. (©Biosphoto/Cordier Sylvain)
Hidrocoria
Por último, está el grupo de plantas que confían sus semillas al agua. Ya se ha citado el ejemplo de algunos cocos que pueden navegar centenares de millas marinas por el océano. Las semillas de otras especies recorren distancias más cortas en cursos de agua o en lagos; por ejemplo, Iris pseudoacorus tiene frutos en forma de banana que, cuando maduran, se abren y liberan cantidades considerables de semillas planas, apiladas unas sobre otras. Estos granos tienen, en el interior de los tegumentos, una bolsa de aire que les permite flotar y, llevadas por la corriente, colonizar otras zonas.
Latencia
El término latencia designa el fenómeno por el cual las semillas, ya maduras y sanas, no germinan durante un determinado periodo o, en cualquier caso, no lo hacen hasta que han pasado, de forma natural o artificial, ciertas fases concretas.
La latencia, cuyas causas físicas o fisiológicas son intrínsecas a la semilla y están programadas de forma genética, impide temporalmente el desarrollo de esta para evitar precisamente que se encuentre en unas condiciones ambientales (agua, oxígeno, luz, temperatura) desfavorables para su germinación.
Es necesario que, a lo largo de la primera fase de germinación, que recibe el nombre de imbibición, haya suficiente agua y calor para que puedan darse un cierto número de reacciones químicas que originan la germinación. Por otro lado, numerosas reacciones son de tipo oxidante y, por tanto, no pueden producirse si no hay oxígeno.
En otras circunstancias, la planta adulta podría encontrarse en dificultades si la semilla germinara demasiado pronto.
Las semillas de las plantas tropicales, típicas de aquellas regiones con clima constante, raramente presentan un estado de latencia; por el contrario, esta se suele manifestar en las especies de los climas templados, que se caracterizan por unos marcados cambios estacionales.

Tapiz de plumas de ara en la orilla del lago de Grand-Lieu, Francia. (©Biosphoto/Balança Erwan)
Condiciones que obstaculizan la germinación
Muchas semillas, cuando llegan a la madurez, tienen un tegumento totalmente impermeable al agua; es el caso de las semillas de Camellia. En la naturaleza, la permeabilidad aumenta con el paso del tiempo: los tegumentos se ablandan y acaban siendo permeables, con mayor o menor rapidez, debido a la acción de los microorganismos presentes en el suelo. También puede pasar al revés: cuando el hombre recolecta el grano, puede aplicarle un tratamiento para modificar la permeabilidad de los tegumentos.
En muchos casos, la rigidez del tegumento es lo que impide el crecimiento del embrión; un ejemplo de ello son los huesos de melocotón. Esta situación puede modificarse por la acción del agua y de los microorganismos, o gracias a la intervención del hombre. En otros casos, el crecimiento del embrión es muy lento y, cuando las semillas se sueltan de las plantas, los embriones todavía no han alcanzado la madurez, de modo que la semilla no puede germinar hasta que el embrión no está listo.
Sin embargo, el caso más corriente es el de un embrión perfectamente desarrollado, cuyo crecimiento está inhibido por unas sustancias químicas contenidas en la semilla, que no podrá germinar hasta que dichas sustancias desaparezcan o hasta que el equilibrio entre estas y las sustancias promotoras se haya inclinado a favor de las segundas.
Puede ocurrir que todas estas condiciones se hallen en una única semilla; el fresno, por ejemplo, tiene tegumentos impermeables, un embrión inmaduro y sustancias inhibidoras en los cotiledones.
Doble latencia
Algunas semillas presentan una característica denominada doble latencia, en cuyo caso la latencia debe finalizar en el momento adecuado para que pueda producirse la germinación. Entre las plantas que presentan doble latencia está el Lilium, los acebos, los tilos y el tejo.
Al inicio se requiere un periodo de tres meses de temperaturas elevadas (entre 20 y 30 °C); durante esta fase se desarrollan las raíces.
Seguidamente se necesitan tres meses más, pero de temperaturas bajas, para interrumpir la latencia de los gérmenes antes de que la planta brote y sobresalga del suelo.
Otras plantas, como Trillium, precisan primero un periodo de tres meses de frío seguido de tres meses de calor, y de nuevo tres meses más de frío antes de germinar.

Flor de lis. (©Biosphoto/Raiser Frederico)