«La información que está dentro de esta primera célula evidentemente le comunica todas las instrucciones de cómo edificarse como un individuo... Me refiero no a una definición de cómo edificar un hombre teórico, sino edificar aquel ser humano en particular que más adelante llamaremos Margarita, o Pablo o Pedro: ya está ahí, pero es tan pequeña que no lo podemos ver».
Testimonio del Dr. Lejeune en el caso de «los embriones congelados de Tennessee»62
Cada vida es un suceso particular que afecta a cada individuo por separado, pero relacionado con otros a través de unas células gaméticas iniciales, el óvulo y el espermatozoide que aportan la información que le dará forma, y que proceden de otros individuos de la misma especie. Estos, a su vez proceden de la unión de óvulos y espermatozoides de otros, y estos de otros anteriores, lo que hace que toda la humanidad comparta un acervo genético común, el genoma propio de nuestra especie, el Homo sapiens. De acuerdo con esto, hay un tronco común y nuestras diferencias biológicas se deben a la identidad genética de cada uno, que se configura en el comienzo de cada vida individual tras la fecundación.
Esto hace que, si bien los planos de ejecución de la obra son comunes para edificar a todos los seres humanos, existan detalles que hacen que el resultado final sea diferente para cada uno. Esos detalles son las variaciones de los genes, los alelos. Pero, debe tenerse en cuenta que no todo es cuestión de genes y que en cada persona a la realidad biológica se unen las dimensiones psicológica y espiritual, no dependientes de los genes.
Siendo esto así, cada ser humano, cada persona, es una realidad singular que comienza su andadura vital cuando se constituye un programa, la información genética que le es propia, pero que labrará en su momento su propia personalidad bajo las influencias del entorno en que vive. Una vida que finalizará con la muerte cuando se suspendan todas las funciones vitales.
Este es el marco en el que se desarrolla la vida humana. Un proceso que con tanta lucidez defendió el Prof. Lejeune como el principal argumento en favor de la vida. En este capítulo vamos a referirnos a los aspectos biológicos de las etapas embrionaria y fetal, tan decisivas en la formación de un nuevo organismo, pero que por la invisibilidad con que se desarrollan tienden a ser ignoradas.
La fecundación. El big-bang de la vida
Los gametos, el óvulo femenino y el espermatozoide masculino, que proceden cada uno de un parental de diferente sexo, poseen 23 cromosomas y una dotación génica simple consistente en unos 21.000 genes. Estas células son necesarias para entre ambas completar la información suficiente para el inicio del desarrollo de una nueva vida.
El encuentro de los dos gametos, constituye el proceso llamado fecundación, que, en la especie humana, de forma natural, tiene lugar en las trompas de Falopio. Estas se encuentran alojadas en el espacio pélvico-visceral de la mujer y conectan la parte alta del útero con los ovarios o gónadas reproductoras femeninas. Desde la pubertad, durante el denominado ciclo menstrual, que tiene lugar aproximadamente cada 28 días, se desprende de uno de los ovarios un óvulo, u ovocito, que pasa a una de las trompas de Falopio. En dicho lugar es donde se produce la fecundación si hasta allí llegan los espermatozoides tras una relación sexual.
El óvulo es una célula esférica, grande e inmóvil, dotada de un gran citoplasma que rodea al núcleo, mientras que los espermatozoides son células pequeñas, piriformes, carentes de citoplasma y altamente móviles debido a la presencia de un largo flagelo en su zona terminal.
Cada espermatozoide posee en la región apical una zona llamada acrosoma, que es una vesícula secretora que juega un papel importante en la fecundación. Producida la entrada de los espermatozoides en el útero estos migran por millares por el tracto genital femenino al encuentro del óvulo. Durante la migración los espermatozoides experimentan una serie de cambios bioquímicos y biofísicos, llamados en su conjunto capacitación, caracterizada por un aumento en la concentración de calcio intracelular y un incremento de fluidez, todo lo cual les prepara para la llamada reacción acrosómica.
La fecundación realmente es un proceso largo que requiere varias horas. Consiste básicamente en la fusión de los dos gametos para dar lugar a la formación del cigoto, pero abarca una serie de pasos que van desde la reacción acrosómica hasta la fusión de los núcleos del óvulo y el espermatozoide.
Antes de proseguir debemos hacer una observación importante con connotaciones bioéticas relacionada con la utilización imprecisa de los términos fecundación y concepción. El primero, también denominado fertilización, no ofrece ninguna duda respecto a su significado biológico, es el proceso de la fusión de los gametos para dar lugar al inicio a la vida. En biología, el término concepción es un sinónimo de fecundación. Sin embargo, de acuerdo con el Dr. Gonzalo Herranz, médico y profesor de la Universidad de Navarra, desde mediados del siglo XX se trató de imponer la idea de que la concepción corresponde a la anidación del embrión en el útero, por el significado etimológico de la palabra, que es retener. Cuando se expandieron las tecnologías de la contracepción, a principios de los años sesenta, los promotores de los métodos anticonceptivos, conscientes de que los métodos dirigidos a interferir con la implantación corrían el riesgo de no recibir una amplia aceptación social, promovieron la idea de declarar éticamente irrelevantes los primeros catorce días del desarrollo embrionario humano, que empieza con la fecundación63. Nada más lejos de la realidad, el verdadero inicio de la vida lo marca la fusión de los gametos, aunque su producto haya de recorrer un camino hasta la anidación en el útero. Desde esta perspectiva la fecundación es el auténtico big-bang de la vida. Es por tanto cuando queda concebida la vida de un nuevo ser humano.
Dada la contaminación que se ha pretendido introducir debe quedar claro que, en adelante, cuando utilicemos la palabra concepción, lo haremos como sinónimo de fecundación, no de anidación, que tendrá lugar días después de la fecundación.
Una vez que un espermatozoide contacta con la cubierta membranosa del óvulo, se produce la reacción acrosómica. Al unirse el espermatozoide con el ligando ZP3, una glicoproteína de la zona pelúcida que rodea y protege al óvulo64, se produce la liberación desde el ápice del espermatozoide, el acrosoma, de unas enzimas hidrolíticas, la hialuronidasa, acrosina y tripsina. Estas contribuyen a alterar la permeabilidad de la membrana del óvulo, facilitan la entrada del núcleo del espermatozoide e inactivan los receptores de su superficie para impedir que otros espermatozoides la atraviesen. Al mismo tiempo se produce desde el espermatozoide la entrada de iones calcio en el óvulo.
Terminada la entrada del pronúcleo espermático en el ambiente citoplásmico del óvulo, queda constituido un ente que posee dos conjuntos de 23 cromosomas, uno aportado por cada gameto. En el óvulo, bajo el estímulo de factores procedentes del espermatozoide se finaliza la meiosis, un proceso generador de los gametos consistente en dos divisiones celulares que tiene lugar en el tejido germinal, y que en la mujer se inició muy pronto, en la etapa embrionaria, y se detuvo en la profase de la primera división hasta la ovulación. De las cuatro células hijas producidas en la meiosis femenina, tres quedan en forma de unos corpúsculos polares, pegados en el exterior de la superficie de la única viable, el óvulo. Tras la eliminación de los corpúsculos, se fundirán los pronúcleos de los gametos masculino y femenino, lo que marca la constitución del cigoto. A partir de este momento es cuando en propiedad empieza una nueva vida.
Antes de seguir conviene hacer otra aclaración. Algunos autores asocian el concepto de vida a las propiedades que tienen las células en general, como la capacidad de reproducción y el metabolismo. Sin embargo, los gametos no tienen capacidad de reproducción, y la vida activa de un espermatozoide o un óvulo se limita a su misión de fusionarse entre sí, de modo tal que si no se culmina este hecho serían eliminados o expulsados como desechos.
Un gameto, como cualquier célula incluso adulta, tiene vida, pero no es un ser vivo. Procede o forma parte de un organismo, pero no es un organismo. Por el contrario, el cigoto posee todo el potencial metabólico, reproductor, de crecimiento y desarrollo. Al constituirse se inicia la vida de un nuevo organismo.
Constituido el cigoto, lo inmediato es su activación metabólica. Esto ocurre tras la liberación de los iones calcio transportados en el acrosoma, desde el lugar de entrada del pronúcleo espermático. Se produce una onda de iones calcio que traza un eje que marca el destino futuro de las regiones del embrión.
De este modo el cigoto representa una realidad celular nueva y distinta, un ente con una organización superior, un organismo. Es la primera realidad corporal del nuevo individuo. Por ello, tiene sentido decir que al constituirse el cigoto se produce el big-bang de una nueva vida. Se pone en marcha el reloj de la vida.
Es preciso recordar que los núcleos de cada gameto se han producido tras pasar por procesos independientes de meiosis durante las gametogénesis materna y paterna. En cada meiosis tiene lugar la recombinación genética y la coorientación de los cromosomas paternos y maternos de cada parental. Por ello, el cigoto posee una combinación cromosómica y génica nueva y distinta a la de los padres de que proceden los gametos.
El cigoto humano posee tres propiedades principales:
- Es un ente genuinamente humano, por tener ADN humano.
- Tiene una identidad genética propia y singular, construida por la adición de genes de sus parentales.
- Es totipotente, ya que esta célula inicial encierra en sí la capacidad de generar todas las células del organismo con sus diferentes especialidades.
Esta célula encierra el programa de desarrollo que inmediatamente se pone en marcha para construir, paso a paso, de forma regular y sin interrupción, el ente que verá la luz cuando alcance el grado de madurez suficiente, lo que, en el hombre, ocurrirá nueve meses después de la fecundación.
De acuerdo con Angelo Serra, profesor emérito de Genética de la Facultad de Medicina de la Universidad Católica del Sacro Cuore de Roma: «el cigoto es el punto exacto en el espacio y el tiempo en el que un individuo humano inicia su propio ciclo vital»65.
Una vez constituido el cigoto comienza el desarrollo embrionario mediante un proceso complejo en el que concurren varios fenómenos.
- La multiplicación de las células por sucesivas mitosis, previa replicación de la información genética contenida en el núcleo de cada célula.
- La diferenciación celular, que va a hacer que las células en proliferación se vayan dirigiendo hacia una futura especialización funcional.
- La organogénesis y morfogénesis, que supone la aparición de estructuras en tres dimensiones constituidas por células organizadas en tejidos que darán lugar a los distintos órganos.
En la dimensión temporal se distinguen dos fases en el desarrollo humano:
- La fase embrionaria, desde el cigoto hasta el final de la séptima semana, y
- La fase fetal, que va desde la octava semana al parto, aproximadamente 36 semanas después de la fecundación.
Todo el proceso de desarrollo está regulado genéticamente en espacio y tiempo. Aunque todas las células reciben la misma información genética del cigoto, se produce una diferenciación de distintos tipos de células, lo que se debe a la expresión diferencial de parte del genoma, permaneciendo inactivos los demás genes en cada una de ellas. Hay por tanto unas instrucciones, un programa de actividades genéticas diferenciales que se pone en marcha tras la fecundación66.
Los avances de la genética del desarrollo y el análisis de los genes que intervienen en cada momento y lugar del embrión durante el desarrollo embrionario, han puesto en evidencia que, en lo que afecta a las decisiones de organización corporal, en los organismos pluricelulares existen dos tipos de genes:
- Los genes estructurales que dan lugar a los tipos de proteínas que determinan la función específica de cada célula, y
- Los genes reguladores, que son jerárquicamente más importantes, por ser los que dan las órdenes de expresión para que se activen los genes estructurales en el momento y en el lugar adecuado.
La actividad de los genes depende del estado de condensación o relajación de la región cromosómica, que como veremos a continuación está determinada por las llamadas modificaciones epigenéticas. Antes de describir su papel en el desarrollo embrionario, describiremos en qué consisten.
Las modificaciones epigenéticas
El término epigenética fue introducido en 1942 por el genetista y embriólogo escocés Conrad H, Waddington (1905-1975) para referirse a la ejecución del fenotipo a partir de las instrucciones potenciales de los genes, bajo el supuesto de que la información de estos ha de recorrer un camino para su expresión en el que pueden ocurrir reajustes, dentro de un rango de variación67.
Bajo determinadas circunstancias, se producen en el ADN de los genes, o en las proteínas a las que está unido en los cromosomas, las llamadas modificaciones epigenéticas que determinan las condiciones fisiológicas de las que depende su expresión. Para que un gen se exprese ha de haber una relajación de la región del cromosoma en que se encuentra, lo que facilitará el acceso de los factores de expresión y de las enzimas que intervienen en la misma.
Las modificaciones epigenéticas pueden afectar al ADN o a unas proteínas llamadas histonas a las que está unido en la conformación de los cromosomas. La principal modificación epigenética que afecta al ADN consiste en la adición de radicales metílicos —CH3— a la base Citosina en los dinucleótidos CpG del ADN de los genes, lo que se llama metilación. La metilación produce una condensación del ADN en donde tiene lugar y conduce al silenciamiento de los genes incluidos en la región afectada.
Las modificaciones que afectan a las histonas son la metilación y otras como la acetilación, la fosforilación y la ubiquitinación. Estas modificaciones afectan a la estructura molecular de los cromosomas, en los llamados nucleosomas —unos complejos de histonas sobre los que gira el ADN—, contribuyendo a dar una mayor o menor compactación a la región afectada, lo que, a su vez, impide o facilita la expresión de los genes. Hoy sabemos que existe un código de histonas, consistente en modificaciones epigenéticas de estas proteínas, que hace más laxa o condensada la fibra de cromatina, permitiendo o no el acceso de los factores y cofactores necesarios para la expresión genética.
Las modificaciones epigenéticas, aun no afectando al ADN en su composición de bases, condicionan la capacidad de expresión de los genes y pueden quedar como una especie de memoria celular, que se puede mantener en el linaje de las células descendientes. Durante el desarrollo embrionario, determinadas regiones de los cromosomas, que contienen genes cuya actividad pasa a ser o deja de ser necesaria, sufrirán las modificaciones que correspondan a las necesidades de su expresión o silenciamiento en las diferentes células del embrión en desarrollo.
Las modificaciones epigenéticas en el embrión temprano
En los vertebrados los núcleos de los gametos masculino y femenino presentan diferentes grados de metilación en sus genomas. Tras la formación del cigoto, y a lo largo de las primeras divisiones de segmentación se produce una serie de cambios muy llamativos en los dos pronúcleos. Las marcas originales de metilación del ADN de ambos pronúcleos se borran, a excepción de unas pocas regiones que corresponden a una impronta genómica paterna o materna de la que hablaremos en el siguiente apartado.
La primera ronda la sufre el pronúcleo paterno que experimenta una desmetilación rápida seguida de la incorporación de histonas maternas y complejos de proteínas. Por su parte, durante las divisiones celulares, el genoma materno sufrirá una pérdida lenta de las regiones metiladas de su ADN68. Al llegar a la mórula —estadio de 8 a 64 células— el genoma estará globalmente hipometilado.
En la transición del estado de mórula a blastocisto comienza de novo una metilación. Las células externas de la mórula darán lugar a la formación del llamado trofoectodermo, mientras que las internas contribuirán principalmente a constituir una masa celular denominada embrioblasto. En el estado de blastocisto, constituido al alcanzar aproximadamente un centenar de células, comienza a introducirse líquido en su interior para formar una cavidad denominada blastocele. En uno de los polos de esta oquedad se concentra el embrioblasto, en el que poco a poco se manifiestan dos capas de células, el epiblasto y el hipoblasto que conforman un disco embrionario bilaminar. Las células del epiblasto darán origen a tres capas de tejido embrionario: ectodermo, mesodermo y endodermo. A ello hay que añadir la capa de células externa, el trofoblasto, que rodea al blastocisto, del que más adelante se derivará la placenta.
Las células del embrioblasto sufren una hipermetilación que afecta a la práctica totalidad del genoma, a excepción de unas zonas, llamadas islas CpG, que permanecen desmetiladas quedando en condiciones de expresarse69 70. Estas regiones son reguladoras de la expresión de los genes. Los estados de metilación o no de diferentes islas CpG en los promotores de diferentes genes, determinan cuáles se activarán o silenciarán en las diferentes células71.
Las células del embrioblasto constituyen las llamadas células madre embrionarias, o más apropiadamente células troncales, y darán origen a los distintos tipos de especialidades celulares de los diferentes tejidos del individuo en formación, por lo que se las califica de pluripotentes. En estas células, tras el borrado programado de las marcas de metilación, se expresan genes nuevos, los epigenotipos propios de cada célula72. Por el contrario, el ADN del trofoblasto permanece hipometilado, lo que es necesario para la correcta implantación en el útero materno73. Los patrones asimétricos de metilación entre el embrioblasto y el trofoblasto persistirán durante el desarrollo embrionario y la placenta, lo que subraya la importancia de las marcas epigenéticas en relación con la funcionalidad e identidad celular.
La impronta genómica materna y paterna
Como hemos señalado, los genomas parentales de los dos gametos presentan diferencias de impronta genómica. Se trata de marcas epigenéticas en dominios específicos del genoma de acuerdo con su origen parental. Las improntas pueden deberse a la metilación de ADN u otros mecanismos.
La impronta se adquiere en función del sexo ya desde el desarrollo embrionario y se materializa durante la gametogénesis de forma diferente en ambos sexos. Muchos experimentos con embriones de mamíferos han demostrado el hecho de la necesidad de que los núcleos gaméticos procedan de dos parentales de diferente sexo, macho y hembra. Experimentos en embriones de ratón demuestran la no viabilidad si han sido constituidos artificialmente con dos núcleos femeninos o dos núcleos masculinos, lo que se debe a las diferencias de impronta de sus genomas74. Los alelos de los genes improntados se mantienen en las células somáticas del adulto, con sus dos estados epigenéticos materno o paterno, de modo que se favorece la expresión de un determinado alelo sobre el otro en función de su origen parental.
Tal como vemos en la Figura 2, la impronta parental materna y paterna, que aportan los gametos y sigue establecida en las células del embrión temprano y en los tejidos somáticos de por vida, solo se resetea en las células primordiales germinales —CPG—, que son el grupo de células del embrión que da lugar al tejido germinal. Una vez que las CPG residen en las crestas genitales, se produce una ronda de desmetilación que borra las marcas improntadas procedentes de los gametos.
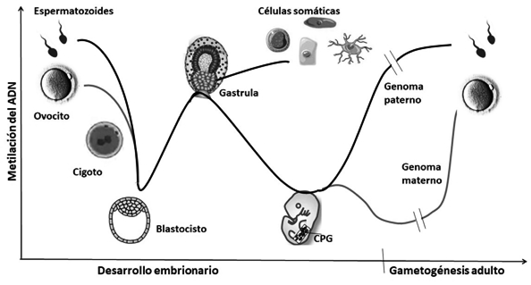
Figura 2. Dinámica de los cambios de metilación en el genoma durante el desarrollo embrionario, tras la formación de los Primordios Celulares Germinales (CPG), en la gametogénesis paterna y materna y su relación con la reprogramación celular en los tejidos somáticos.
La gametogénesis femenina, ovogénesis, comienza durante los primeros meses del desarrollo fetal femenino, pero no se completa. Las células que inician la meiosis para generar los gametos detienen el proceso en la profase de la primera división meiótica, en el estado de dictiotena, y así permanecerán hasta llegar a la madurez sexual. La impronta materna se produce al completar la ovogénesis, de manera asincrónica, cuando se reanuda la meiosis para dar lugar a un óvulo en cada ciclo menstrual75.
Por el lado masculino, las CPG de la línea germinal adquieren la impronta en el estado embrionario-fetal con la metilación de las regiones específicas de la impronta paterna, mucho antes de la gametogénesis, o espermatogénesis. De este modo, hay un desfase en la adquisición de la impronta dependiendo del sexo del individuo en desarrollo.
Las modificaciones epigenéticas y la diferenciación celular
Tras el estado de blastocisto y cuando el embrión se implanta da comienzo la fase de gástrula. Se produce una hipermetilación que da lugar a una represión global del conjunto de genes menos un mínimo en cada célula.
Esto significa que en el desarrollo embrionario, aunque el genoma individual se transmite completo de célula en célula, en cada una de ellas empieza a variar el epigenoma, es decir, la parte del genoma que se expresa, contribuyendo de esta forma a la diferenciación celular.
El mecanismo que lo hace posible es el programa de modificaciones epigenéticas que sigue unas instrucciones perfectamente reguladas para hacer posible la especialización de las células y tejidos. Ocasionalmente puede ocurrir que, bajo la influencia de factores ambientales, durante el desarrollo embrionario, o también en tejidos en la fase adulta, se produzcan modificaciones epigenéticas no programadas en algunas regiones de la cromatina, lo que podría afectar a la expresión de algunos genes. Estas alteraciones pueden deberse a factores fisiológicos y ambientales no controlados y puede tener consecuencias para la salud.
Las modificaciones epigenéticas no programadas que afecten a los tejidos somáticos no se heredarán y afectarán al embrión o al feto en formación. Por el contrario, las que afectasen al tejido germinal podrían pasar a la siguiente generación y sus efectos podrían llegar incluso a la segunda generación filial, en el caso de no ser borradas a su paso por la embriogénesis tras la fecundación. Pero no más allá de dos generaciones.
El programa de modificaciones epigenéticas durante el desarrollo embrionario no afecta especialmente al trofoblasto, desmetilado para la correcta implantación en el útero materno e intercambio de nutrientes76.
Debe de quedar claro de todos modos que las modificaciones epigenéticas programadas que se producen en el embrión a partir de la gastrulación no están determinadas desde el ambiente materno, sino que siguen las instrucciones del desarrollo autónomo del propio embrión.
En síntesis, el desarrollo embrionario obedece a un programa perfectamente coordinado de actividades genéticas, regulado en espacio y tiempo. Es continuo, ya que de una etapa se pasa a la siguiente sin solución de continuidad, es gradual, ya que a medida que pasa el tiempo surgen rutas de especialización de las células que lo integran, y es complejo, por la diversidad creciente de tipos de células con misiones diferentes.
Volvamos ahora al principio y sigamos las diferentes fases del desarrollo embrionario y fetal humano.
El desarrollo embrionario desde el cigoto
Terminada la fecundación, el embrión es ya un individuo humano que ha comenzado su existencia y en el que se cumplen todas las condiciones necesarias y suficientes para alcanzar de forma autónoma todo el potencial para el que está genéticamente equipado.
Aproximadamente a las 24 horas de la constitución del cigoto habrá tenido lugar la replicación del ADN y, tras ello, el reparto de las copias del genoma a las dos células hijas mediante el mecanismo de la mitosis. Las dos células hijas volverán a dividirse, lo mismo que sus células descendientes y así sucesivamente. Previo a cada división celular hay una replicación del ADN, de modo que las células hijas reciben siempre una copia exacta de la información genética que se constituyó en la fecundación. La etapa embrionaria se caracteriza por la proliferación celular a base de mitosis.
Los linajes celulares se van preparando para sus futuros papeles diferenciales, prácticamente desde la primera división de segmentación77. La iniciación de la transcripción —expresión de los genes— propios del embrión, se conoce como la Activación del Genoma Cigótico —AGC—, y ocurre en varias olas sucesivas. La AGC comienza en el cigoto y continúa con importantes ráfagas de transcripción de novo en las etapas de 2 y 4 células. Pronto se degradarán los productos de la transcripción de origen materno presentes en el óvulo.
Mientras aumenta el número de células, el embrión va descendiendo por la trompa de Falopio hacia la pared de la parte inferior del útero, donde anidará. Aproximadamente a los tres días de la fecundación el embrión tiene aspecto de una mora, por lo que se le denomina mórula. En su proceso de crecimiento pasa por 8, 16 y 32, células entre las que ya se expresan genes del propio embrión78. Al mismo tiempo, a medida que crece el número de células aumenta la adhesión entre ellas.
Al cabo de cinco días el embrión alcanza el estado de blastocisto, que ya posee entre 64 y 128 células. Llegado este momento, comienza a introducirse líquido en su interior para formar el blastocele.
A pesar de la apariencia más o menos regular en forma de esfera de la mórula o del posterior blastocisto, este embrión es mucho más que un conglomerado de células, precisamente por las diferencias de expresión de los genes en cada una de ellas.
Las dos células de la primera división de segmentación tienen una asimetría en la concentración de iones calcio, lo que hace que, si bien poseen una copia idéntica del genoma del cigoto, no sean iguales en sus contenidos citoplásmicos. La que más calcio tiene se divide antes, originando un embrión tricelular. Las células ricas en calcio mantienen la pluripotencialidad y sus descendientes se quedarán en el interior para dar lugar al embrioblasto. La célula con menor concentración en calcio y sus descendientes formarán el trofoectoblasto. La Dra. Magdalena Zernicka-Goetz demostró las diferencias de destino de las células resultantes de la primera división de segmentación del cigoto, en un experimento que consistió en la aplicación de unos fluorocromos79 de distintos colores. Esto permitió visualizar el destino de las células hijas durante el desarrollo embrionario. De acuerdo con la expresión de dicha investigadora: «en la primera división celular ya existe una memoria de nuestra vida»80.
La especificación del linaje de cada célula está regulada genéticamente por los productos de la transcripción de unos genes reguladores. Los primeros genes reguladores que se activan son Oct-4, Nanog y Sox-2, que son esenciales para establecer las diferencias en la potencialidad de las células del embrión temprano. Mediante su interacción cooperativa, se impulsa la expresión de otros genes como requisito necesario para la diferenciación81 82.
Se conocen varios modos diferentes por los que en las células se estimula la expresión de genes específicos según su posición en el embrión durante el desarrollo. Las células se comunican por señales. Los genes que se van expresando determinan la síntesis de unas proteínas que actúan como una señal que generalmente es reconocida por las células vecinas a través de receptores de la pared celular. A su vez, estas señales inducen una cascada de actividades de otros genes en el interior de cada célula de acuerdo con su posición en el embrión.
Las señales intercelulares durante el desarrollo embrionario se materializan bien por contacto físico directo, o a distancia, ya que las moléculas de señalización, proteínas, o también ARN de interferencia o ARN no codificante, son secretadas al exterior y pueden llegar a otras células situadas desde solo uno o dos diámetros de la célula emisora hasta el de 50 células o más. De este modo se establece un diálogo entre unas células y otras que determina la activación o silenciamiento de genes diferentes en cada una de ellas. La expresión o silenciamiento se produce de acuerdo con las modificaciones epigenéticas de las regiones del genoma en cada célula, de acuerdo con el momento del desarrollo y su posición en el embrión. De este modo, cada célula adquiere un epigenoma propio de acuerdo con su posición en el embrión y momento del desarrollo, mediante la interpretación del mensaje que le llega del resto del embrión.
En los embriones femeninos de los mamíferos, que poseen dos cromosomas X, a diferencia de los masculinos que tienen solo uno, tiene lugar un mecanismo muy peculiar, llamado lionización, que consiste en la inactivación en cada célula de uno de los dos cromosomas X, el paterno o el materno al azar, ya desde las primeras divisiones celulares. En la anulación de uno de los cromosomas X intervienen unas moléculas de ARN de interferencia. Al quedar inactivo uno de los dos cromosomas X, se produce una compensación de dosis génica en las hembras, respecto a los genes del único cromosoma X presente en el genoma de los machos. Esta inactivación de un cromosoma X al azar se mantiene en el linaje celular de por vida, de modo que las hembras de mamífero presentan un mosaicismo respecto a los genes paternos o maternos, que se expresan en sus diferentes órganos y tejidos.
Llegado al estado de blastocisto, hacia el final de la primera semana del desarrollo, el embrión anida en la mucosa uterina a través de las células del trofoectoblasto que recubren el polo más próximo al embrioblasto. Hacia el octavo día después de la fecundación se organiza la cavidad amniótica, que se constituye en el ambiente protector del embrión a partir de células del ectodermo y el endodermo primitivo.
Tras la implantación se produce una relación perfectamente armoniosa entre el embrión y las células del endometrio del útero que le da cobijo. El embrión, que posee un perfil genético de histocompatibilidad diferente al de la madre, no es rechazado como un cuerpo extraño, sino acogido y protegido como propio. Este hecho tiene su fundamento en una modificación molecular de los mecanismos inmunológicos maternos a favor del embrión, lo que no debe interpretarse con la infundada idea de que el embrión o el feto se convierte en un órgano más de la madre, sino como el fruto de una adaptación fisiológica para favorecer la protección del feto.
Hacia el décimo día tiene lugar el desarrollo del corion, que se constituye a partir de las células del trofoectoblasto junto con células del mesodermo del embrioblasto. El amnios es transparente y contiene el líquido amniótico que protegerá al embrión hasta el parto. El líquido amniótico es un líquido acuoso que rodea al feto, amortigua los golpes, proporciona flotabilidad y le permite moverse libremente dentro del espacio del útero. El corion se convierte en la parte fetal de la placenta que protegerá al propio feto a partir de ese momento.
Transcurridos catorce días desde la fecundación el embrión ha seguido creciendo y ha aumentado su asimetría morfológica y funcional. Tras la anidación, hacia la tercera semana, comienza la gastrulación, una fase que se caracteriza por la aparición de la llamada estría primitiva, una banda celular engrosada situada en la región central posterior del embrioblasto superpuesta a los tres estratos celulares: el ectodermo, el mesodermo —ya claramente definido—, y el endodermo. A partir de estas capas celulares primitivas, merced al proceso de emisión de señales entre las células, se empezarán a diferenciar los tejidos. Con la gastrulación queda definido el plan general del cuerpo y da comienzo la organogénesis.
Hacia la quinta semana el embrión, que puede haber alcanzado los seis o más millones de células, alcanza un milímetro de longitud. Queda iniciado el proceso de la morfogénesis. En este momento, se encuentra claramente orientado el desarrollo del cerebro, el corazón, los pulmones y esbozos de otros órganos. El corazón late a un ritmo de unas 113 veces por minuto.
A las seis semanas se observa un intenso crecimiento del cerebro a partir del tubo neural. Emergen los hemisferios cerebrales y la cabeza llega a medir un tercio del total de la longitud corporal. El embrión empieza a hacer movimientos espontáneos y reflejos. Al mismo tiempo ya están presentes los bronquios en el aparato respiratorio, aún no conectados a la tráquea. Simultáneamente aparecen los riñones. Se empiezan a formar las orejas. Se desarrolla el sistema inmunológico y aparecen los linfocitos.
A las siete semanas, el corazón posee ya las cuatro cámaras: dos aurículas y dos ventrículos, y late a un ritmo de 167 latidos por minuto. La actividad eléctrica cardiaca es similar a la de un adulto. Poco después aparece la retina pigmentaria en los ojos. Los dedos de las manos ya están separados y los de los pies aparecen unidos por su base. Las rodillas desarrollan sus articulaciones.
Son necesarias alrededor de siete semanas para alcanzar el punto crítico del comienzo de la fase fetal, caracterizado por la formación de todos los tipos de tejidos y órganos necesarios que se van completando durante la etapa fetal, hasta el nacimiento.
El período de la cuarta a la novena semana constituye probablemente la etapa más crítica de expresión del genoma para el desarrollo de la mayoría de los órganos. Aproximadamente la mitad de los genes humanos se expresan durante ese período. La perfecta armonía en el desarrollo gradual del embrión solo se vería alterada de haber algún gen regulador o estructural defectuoso de los que intervienen en el desarrollo, o de mediar algún factor externo perturbador. Las mutaciones pueden ser heredadas o producidas de forma espontánea o inducida. Cuando hay un gen defectuoso o cuya expresión se ve alterada por una modificación epigenética, sea o no programada, o por efecto de algún agente externo, químico o físico, como drogas, alcohol, radiaciones, etc., puede verse afectado el desarrollo de un tejido o un órgano, dando lugar a malformaciones en el feto que incluso pueden dar origen a un aborto espontáneo o una malformación. Este tipo de agentes se denominan teratógenos. Las alteraciones genéticas y teratogénicas pueden ser la causa de cerca del 15% de abortos espontáneos de origen desconocido.
Recordaremos al respecto el caso de los efectos teratogénicos de la talidomida, un analgésico producido en Alemania a finales de los años cincuenta y comercializado en muchos países europeos, diseñado para paliar las náuseas y molestias de las mujeres embarazadas. La talidomida fue la causa de miles de nacimientos en Europa de bebés con malformaciones y terribles focomelias debidas a fallos en el desarrollo del tejido óseo y muscular. En 1963, tras descubrir la causa de estas minusvalías, se retiró el producto de las farmacias.
Un apartado especial de los genes reguladores que intervienen en el desarrollo lo constituyen los genes llamados homeóticos, de los que dependen las decisiones de organización de una parte determinada del cuerpo, anterior o posterior, dorsal o ventral, cabeza, tórax, extremidades, ojos, etc. Las mutaciones de estos genes repercuten especialmente en el patrón de desarrollo y pueden generar modificaciones fenotípicas conocidas como transformaciones homeóticas.
Este tipo de genes reguladores se han mantenido con un extraordinario grado de conservación en diferentes líneas evolutivas. Así, por ejemplo, existe una gran similitud entre los genes homeóticos Hox, de los mamíferos, los de los sistemas HOM-C del coleóptero Tribolium castaneum y los llamados ANT-C y BX-C de Drosophila melanogaster y en general en todos los animales. Es interesante el hecho de que estos genes presenten sus subunidades génicas dispuestas en un orden lineal correspondiente al de su expresión espacial y corporal. Se ha propuesto que los genes homeóticos Hom y Hox aparecieron en la explosión de formas de vida hace unos 570 millones de años, durante el Cámbrico, y se han mantenido desde entonces en las líneas evolutivas derivadas de un ancestro común. Este tipo de genes son la causa de las diferentes arquitecturas corporales que diferencia a los grupos de animales, habiendo contribuido a la biodiversidad a lo largo de la evolución, por variaciones debidas a mutaciones con consecuencias en la organización espacio-temporal del esquema corporal. Est explica el fenómeno conocido como macroevolución.
Los genes Hox tienen diferentes papeles durante el desarrollo de los vertebrados. En el genoma humano, hay cuatro grupos de genes Hox, con localización en diferentes cromosomas. Estos genes se expresan pronto para la constitución del sistema nervioso central, tras la formación de la placa neural durante la gastrulación.
Muchas enfermedades congénitas cardiacas, de los sistemas nervioso, esquelético, digestivo, etc. se deben a mutaciones o fallos de expresión de este tipo de genes durante el momento crítico de formación de los órganos correspondientes, lo que puede dar lugar a las discapacidades o disfunciones que se revelarán más adelante. Un buen ejemplo de la participación de los genes homeóticos en la morfogénesis de determinados órganos lo ofrece el gen PAX6, que interviene en el desarrollo ocular. Una mutación semidominante en este gen puede determinar su pérdida de función y conducir a anomalías en la retina, el cristalino, la córnea o la aniridia, que consiste en la ausencia o reducción del iris.
Nuevas tecnologías aplicadas al estudio del desarrollo embrionario
Al conjunto de conocimientos derivados de los estudios de la biología molecular, la biología celular y la embriología, se han añadido recientemente una serie de nuevas tecnologías que permiten abundar en el conocimiento del desarrollo embrionario. Estas técnicas son la secuenciación del ARN de una sola célula —scRNA-seq—; la preparación de embriones de ratón fuera del útero para su observación in vivo; y la microscopía confocal unida a la utilización del marcado con proteínas fluorescentes para la interpretación de las imágenes por computación, etc.
Así, la técnica denominada scRNA-seq facilita secuenciar el ARN extraído de las células de las diferentes partes del embrión en diferentes momentos del desarrollo, lo que ha permitido abordar lo que se ha dado en llamar el Atlas del Desarrollo Celular Humano —HDCA—83. La técnica permite conocer cómo se generan paso a paso los diferentes tipos de órganos en la fase fetal o en la base de los tejidos adultos. La scRNA-seq es ya lo suficientemente fiable, precisa y sensible como para habilitar la detección incluso de las transcripciones de bajo nivel en las células84.
El HDCA ha abierto el camino al conocimiento de una amplia gama de cuestiones biológicas tales como el origen de las malformaciones, el envejecimiento, el cáncer y la reproducción. Además, permitirá cancelar cualquier especulación sobre la naturaleza biológica y el valor de la vida humana, al desvelar cómo crece de forma regular, gradual y continua un ser humano desde la fecundación.
En segundo lugar, las mejoras técnicas de la preparación de embriones vivos de ratón fuera del útero, junto con mejoras en su cultivo in vitro y de los métodos de su montaje para la observación en un microscopio confocal, han permitido reconstruir los detalles de la evolución de las diferentes partes del embrión a lo largo del tiempo.
Gracias a esto, se ha podido visualizar la morfogénesis embrionaria en embriones de ratón, con un detalle y una resolución extraordinarios y sin precedentes, lo que ha permitido obtener imágenes del desarrollo embrionario en 4 dimensiones85.
Por qué un embrión es más que un cúmulo de células
Si consultamos los diccionarios científicos especializados encontraremos en todos ellos definiciones muy claras sobre lo que es un embrión. Así, en un Diccionario de Biología muy popular se define el embrión como «el animal en proceso de desarrollo a partir del huevo fecundado»86, a lo que Eldra Solomon y otros autores, en su tratado de biología, añaden que el embrión «es el organismo multicelular en la fase inicial de la vida, antes de que salga del huevo, semilla o cuerpo materno, definición que aplicada a la especie humana se extiende hasta el término del segundo mes del desarrollo, después de lo cual se denomina feto»87. Reparemos en el significado de las palabras de esta definición cuando se señala que el embrión es un organismo multicelular, y no un agregado multicelular. Algo que también se reitera en el Glosario de Genética y Citogenética, de Rigomar Rieger y otros autores, que definen el embrión de los animales como «el organismo joven que surge de la célula huevo fecundada»88.
Es significativo que todas estas definiciones sean anteriores al desarrollo de las técnicas de fecundación in vitro, por lo que no están contaminadas por la corriente utilitarista que ha convertido a los embriones en objetos de experimentación.
Pero es muy importante resaltar que un embrión es un todo integrado en cada momento y que el programa de desarrollo por el que atraviesa se plasma en una perfecta coordinación e interdependencia a los niveles celular y molecular, debido al programa de actividades genéticas a lo largo del tiempo y del espacio que en él tiene lugar.
En ningún momento se puede considerar al embrión como un mero conglomerado o acumulo de células, término que según el diccionario de la RAE se aplica a la acción y efecto de unir cosas sin orden. El embrión no es una suma de células sino un todo estructural y funcionalmente integrado. La palabra que mejor define este todo integrado es organismo, del que el diccionario de la RAE dice es un ser viviente. Lo que se desarrolla de forma coordinada, continua y gradual es un organismo, un ser humano en sus primeras etapas de desarrollo. Aunque en un momento dado no se aprecien diferencias externas, conforme vaya creciendo en tamaño y vayan produciéndose más células, aumentará su complejidad sin cambiar la identidad genética de partida. Aunque todas las células de un embrión tengan la misma información y, a pesar de la apariencia, en cada célula y momento del desarrollo solo se expresa la parte del genoma necesaria de acuerdo con su contribución al conjunto del organismo. El embrión que se desarrolla es siempre el mismo organismo y si es humano es indiscutiblemente un ser humano.
Desde la perspectiva espacial y temporal, tan incorrecto es considerar la vida humana solo en relación a una de sus etapas, minimizando o negando la realidad de la existencia en las demás, como pretender reducirlo a un simple aglomerado de partes inconexas. Cada vida es un todo integrado en sus dos dimensiones: espacial y temporal.
En el caso humano, la complejidad del desarrollo va en aumento hasta llegar a un organismo con billones de células especializadas y repartidas en unos 220 tipos de especialidades celulares. Es, desde la fecundación, el mismo organismo, que vive su vida de forma personal hasta la pérdida de sus funciones vitales.
El embrión: ¿ente biológico o entelequia filosófica?
Tras lo señalado no debe quedar ninguna duda de que para la biología un embrión es un ente vivo en las primeras etapas de su desarrollo desde la fecundación. Sin embargo, desde posiciones más filosóficas que biológicas se plantean algunas cuestiones que conviene aclarar.
En primer lugar, se habla del estatus del embrión desde varias perspectivas diferentes que corresponden a distintas aproximaciones: biológica, ontológica, ética y jurídica. Se trata de analizar si el embrión humano debe considerarse un ente vivo de la especie, si ha de concedérsele la consideración de persona, si tiene un valor moral intrínseco y en qué medida ha de ser protegido. De todos estos niveles, el biológico, lo que dice la ciencia sobre el embrión humano, es fundamental, pues difícilmente podríamos conceder el concepto de persona con la dignidad que le corresponde y el reconocimiento de su valor ético a algo de lo que se ignorase su naturaleza. Si hubiese que establecer una prioridad, la última palabra la tendría la aproximación jurídica, que, en función de la naturaleza biológica, la dignidad y el valor ético que se reconociese al embrión, debería servir para establecer las normas o leyes de protección que le correspondieran, con sus deberes y obligaciones.
A pesar de ello, el debate no ha seguido este orden, y especialmente desde una perspectiva utilitarista, la discusión se ha embarrado al llevarla a un terreno más especulativo o deliberativo que real ignorando la naturaleza biológica y el valor intrínseco de la vida humana en su etapa más vulnerable.
Así, el Prof. Diego Gracia89 centra el debate sobre el valor humano del embrión en desarrollo en el concepto filosófico de constitución y se plantea cuándo el no nacido tiene entidad constitutiva, es decir cuándo se le debe atribuir el estatus de persona. Cuando se habla de constitución, de acuerdo con la acepción del diccionario de la RAE, estamos hablando del conjunto de caracteres específicos de algo. Es decir, el conjunto de notas que caracterizan un todo. Seguidor del filósofo y teólogo Xavier Zubiri (1898-1983), el Dr. Gracia, se plantea el momento en que a lo largo del desarrollo la estructura en crecimiento adquiere las notas que le confieren suficiencia constitucional. El filósofo Xavier Zubiri señalaba que para que haya suficiencia constitucional debe haber sustantividad y Diego Gracia90 no reconoce en la información genética constituida tras la fecundación la suficiencia constitucional, que según opina se completa con aportaciones de la madre. Es decir, plantea que un organismo en desarrollo en el que van surgiendo nuevos caracteres, nuevas notas que lo conforman, no adquiriría estatus de ser vivo hasta que se manifestasen todos los elementos constitutivos de su naturaleza. Como tras la anidación, el embrión recibe el cobijo necesario para ir avanzando en su desarrollo bajo el aporte de las condiciones físicas y los metabolitos necesarios por parte de la madre, a juicio del Dr. Gracia, el embrión carecería de suficiencia constitucional hasta que se completase tal aporte. Según esto, todo el camino recorrido por el embrión no merecería el tratamiento de ser humano hasta al menos la octava semana, quedando justificada la negación de cualquier derecho en esta crucial etapa del desarrollo de la vida humana.
Dados los avances de la biología celular y molecular y de la genética del desarrollo, hoy sabemos que cada célula del embrión cumple un papel en interacción con el conjunto. Pero el conjunto es una unidad de desarrollo en sí misma que se autoconstruye con el programa genético que quedó fijado en el cigoto. Todas las células de un embrión temprano contienen la misma información de partida, que incluso utilizan solo parcialmente. Sin embargo, cada célula del embrión temprano, lejos de ser un elemento autónomo, contribuye de forma diferencial al proceso del desarrollo del todo, que, además, debe observarse con perspectiva temporal. El desarrollo es un proceso dinámico y continuo y, si bien poco a poco van aflorando nuevas características, sería arbitrario determinar un momento concreto para decidir que ya posee suficientes notas como para reconocerle el estatus de persona. ¿Cómo decidir ese momento si la realidad es la misma que ya existía desde la fecundación y seguirá siéndolo hasta el final de su vida?, y ¿cómo atribuir al ambiente materno la incorporación de unas notas que no es que no sean propias del embrión, sino que no ha transcurrido el tiempo necesario para que se manifiesten? El ambiente materno del que depende el desarrollo embrionario no constituye un elemento formal del embrión, aunque sea necesario para su desarrollo, como no lo es el aire o la alimentación del recién nacido o del adulto.
Del mismo modo, el genetista Carlos Alonso Bedate (1935-2020)91, profesor de Investigación del C.S.I.C., cuestionó la condición de ser humano al embrión antes de la octava semana, no por carecer de suficiencia constitucional, sino por «la dependencia del genoma del embrión del genoma de la madre, sin cuya dependencia devendrían molas hidatiformes».
Sin embargo, hay que insistir en la capacidad potencial del embrión para desarrollarse normalmente de no existir ninguna alteración en el programa genético de partida y de no mediar una perturbación que lo alterase una vez iniciado su desarrollo. Este, por supuesto pasa por la implantación y protección en el seno materno, pero ello no implica que le falte información genética que vaya a ser aportada por la madre, sino en todo caso que necesita un entorno ambiental fisiológico y molecular adecuado para que se despliegue todo el programa genético que posee desde la fecundación. Tras la implantación y una vez establecida la placenta que media entre el embrión y el claustro materno, pero que se genera a partir de las células del trofoblasto del propio embrión, este prosigue su impulso morfogenético dependiendo de su equipamiento genético, en las condiciones óptimas de desarrollo.
En segundo lugar, habría que precisar algo sobre la posibilidad de que surjan estructuras celulares alteradas no equivalentes a los embriones. Este es el caso de los llamados partenotes y las molas hidatiformes, que desde el punto de vista biológico podrían homologarse al producto de la partenogénesis que existe como un mecanismo de reproducción en muchas especies de plantas y animales, pero no en el hombre.
En algunas especies animales, bajo la influencia de un estímulo químico o físico, el óvulo no fecundado se comporta como si lo hubiese sido, y da lugar al partenote, que sigue un desarrollo semejante a un embrión. Se han descrito incluso casos en tiburones y ratones, en que las hembras no fecundadas pueden dar lugar a partenotes que pueden madurar y llegar a adultos92. Sin embargo, cuando esto ocurre en los mamíferos, las estructuras resultantes son inviables y no sobreviven hasta el nacimiento93. Esto evidencia, una vez más, la importancia del programa genético propio y equilibrado para el normal desarrollo de los embriones. No hay pruebas de que los ocasionales partenotes humanos presenten una actividad genética coordinada capaz de generar un patrón completo de desarrollo.
Un caso semejante es el de la aparición de una mola hidatiforme. Se trata de una estructura de aspecto tumoral, que se produce como consecuencia de la entrada de un núcleo espermático en un óvulo que ha perdido su propio núcleo, quedando por tanto solo el núcleo paterno94. Esta alteración, tiene una frecuencia de 1 en 1000 embarazos y los óvulos así formados son incapaces de desarrollarse por problemas en la constitución del trofoblasto y la placenta, por lo que a esta alteración también se la suele llamar enfermedad trofoblástica gestacional.
La inviabilidad de los partenotes y las molas hidatiformes ponen de relieve la importancia del desequilibrio genético que contienen por carecer de la información genética completa de dos núcleos, con una impronta materna y paterna diferente y complementaria. Evidentemente estas estructuras difícilmente serían viables.
Es cierto que también puede haber embriones portadores de alteraciones cromosómicas inviables, como muchas aneuploidías o deleciones cromosómicas importantes, responsables de cerca del 50% de los abortos espontáneos. Pero a diferencia de las molas hidatiformes y los partenotes, se trata de embriones humanos que reúnen la información de dos gametos con la información de dos parentales, como es norma en la reproducción sexual. Por supuesto, se trata de embriones humanos, aunque lleven alteraciones cromosómicas. Entre este tipo de casos se encuentran los embriones portadores de aberraciones cromosómicas, las poliploidías, aneuploidías y las deleciones de regiones cromosómicas, como la trisomía 21 que determina el síndrome de Down, la deficiencia de un cromosoma X en la mujer (45, X0), causante del síndrome de Turner, y muchas otras cromosomopatías viables o no viables.
Otra idea carente de sentido biológico es la de quienes sostienen la hipótesis de la animación retardada, que hace hincapié en la importancia de la anidación como fundamental para la aparición de la vida humana.
La anidación no supone ningún cambio en la esencia del embrión, que, tras la implantación en el útero materno, sigue su transformación en relación con su entorno, pero en dependencia de su propia información genética. Lo que ocurre, tras la anidación, es que se acentúa su relación y dependencia del ambiente materno hasta el extremo de que si el embrión no anida se detiene el desarrollo y muere, pero no por sus carencias constitucionales, sino por la falta de un ambiente adecuado que le proporcione el oxígeno y los nutrientes necesarios para el mantenimiento de la vida. Por ello, como bien decía la Dra. Vila-Coro: «La anidación en el útero materno no añade ni quita nada a la nueva vida en sí misma; lo que hace es suministrarle las condiciones ambientales óptimas para su desarrollo»95.
De todo lo anterior se pueden colegir dos conclusiones:
- Que el embrión —en virtud de la continuidad y autonomía de su desarrollo desde la fecundación y antes o después de implantarse en el útero—, es siempre la misma vida, el mismo ser humano, que poco a poco se desarrolla hasta el nacimiento.
- Que el embrión —desde la fecundación hasta la octava semana—, y el feto —a partir de la octava semana—, son las primeras fases del desarrollo de un nuevo ser humano y en el claustro materno no forman parte de la sustantividad ni de ningún órgano de la madre, aunque dependa de esta para su propio desarrollo.
El absurdo concepto de preembrión
Como veremos en el siguiente apartado hay quienes sostienen que la condición humana del no nacido depende de su individualidad. Es decir que no se ha de conceder la condición de ser humano, y aun de vida humana, a algo que no tiene garantizada la unicidad. Es decir, que se trata de un único individuo. Para los defensores de esta idea, la unicidad no queda garantizada hasta la anidación96. Esta idea fue planteada por el filósofo y teólogo australiano Norman Ford, sacerdote salesiano y profesor de la Universidad de Melbourne, que planteaba el problema de la gemelación como dificultad fundamental para que exista un ser humano individual, al señalar que «la potencialidad de la división gemelar monocigótica es incompatible con el estatus personal»97. Esta forma de pensar dio paso a la idea de la bióloga inglesa Jeanne McLaren de considerar que hasta el decimocuarto día después de la concepción no debe hablarse de embrión, sino de preembrión98, término que se utilizó por primera vez en el informe Warnock, a resultas del cual fue aprobada la utilización de los embriones en investigación por el Parlamento del Reino Unido en 1984. Más tarde, la Dra. McLaren confesó su equivocación.
El término preembrión no tiene cabida en ningún tratado de biología o de embriología, y solo se aplica con un sentido utilitario en normas o textos de carácter jurídico. Es un término totalmente incorrecto que no aparece en la literatura científica sino únicamente en los textos jurídicos de determinados países, como ocurre en España, con la Ley de Técnicas de Reproducción Asistida de 1988 y sus sucesivas reformas de 2003, 2006 y 2007.
Lo cierto es que minimizar el significado del embrión durante las primeras etapas de la vida humana, ignorando de paso la identidad genética desde la fecundación, solo puede obedecer a una intención. La de justificar la utilización de los embriones obtenidos por fecundación in vitro con fines experimentales.
Al margen de la intencionalidad, la realidad es que la gemelación es un suceso accidental que tiene una probabilidad inferior al 0,2%, que lo único que demuestra es que la individualidad genética embrionaria inicial no implica indivisibilidad. Además, existen serias dudas sobre el límite temporal del suceso de la gemelación, que algunos pretenden extender hasta la anidación. Lo cierto es que, desde la fecundación hay un ente real, una realidad biológica que seguirá siendo única en el 99,8% de los casos. Como bien señala el ginecólogo español José Antonio Usandizaga: «No ha de confundirse la individualidad con la indivisibilidad, ya que aunque durante los primeros días del desarrollo del embrión exista la posibilidad de una división gemelar, eso no quiere decir que dicho embrión carezca de individualidad»99.
Profundizaremos entonces en la incidencia de la gemelación por su protagonismo en el marco de la bioética.
La partición y segregación de partes del embrión para reiniciar el camino del desarrollo como si se tratara de entes diferentes, es un raro accidente que puede tener lugar al principio del desarrollo. Es necesario distinguir entre dos tipos de gemelos: los dicigóticos o fraternos, y los monocigóticos o idénticos. Los gemelos dicigóticos no se producen por la segregación de un embrión inicial, sino por una fecundación doble —dos espermatozoides que fecundan a otros tantos óvulos o al óvulo y a un corpúsculo polar—. Los gemelos idénticos o monocigóticos se originan por la segmentación de un embrión único y constituyen la única posibilidad real de que dos personas distintas posean la misma identidad genética.
Probablemente de forma errónea se ha mantenido durante años la creencia de que durante las etapas tempranas del desarrollo los blastómeros conservan su totipotencialidad, por lo que, si se subdividiese accidentalmente el embrión, las partes separadas podrían proseguir un desarrollo embrionario independiente. De ahí la idea de que mientras no exista individualidad no cabe hablar de vida humana. Es decir, se exige la unicidad como requisito para considerar que estamos ante un nuevo individuo de la especie. Sin embargo, la suposición de que la segmentación accidental del embrión puede suceder hasta el estado de blastocisto, carece de demostración experimental.
El Dr. Gonzalo Herranz, discute los modelos poco probados, y más teóricos que basados en demostración experimental, que dicen que los gemelos monocigóticos se originan por una partición en fase avanzada del desarrollo embrionario100 101. En su argumentación, el Dr. Herranz avanza una nueva teoría según la cual el accidente de la partición de un embrión sucedería pronto, poco después de finalizar el proceso de la fecundación. Según esta idea, el gemelismo podría tener lugar en la primera división de segmentación, que da paso a la transición al estadio de dos células, que al separarse podrían dar lugar a dos gemelos genéticamente idénticos. Cada uno generaría su propio trofoblasto del que se derivarían sus propias membranas protectoras. Es decir, serían gemelos cada uno con su propio corion —gemelos dicoriónicos—, y su propia placenta —diamnióticos—, como también ocurre normalmente con los gemelos dicigóticos. De hecho, los gemelos dicoriónicos y diamnióticos suponen entre el 25% y el 30% de los gemelos monocigóticos.
Por otra parte, los embriones gemelares independientes, que comparten corion y amnios, podrían originarse no por escisión en dos de un embrión en fases más avanzadas de su desarrollo, sino por la fusión de las membranas que envuelven a los embriones monocigóticos separados desde el principio, tras la primera división de segmentación.
Esta explicación de la gemelación es trascendental para el debate bioético, pues en realidad ya no cabría esperar a la anidación o a la formación de la estría primitiva como momento en que se garantizaría la unicidad embrionaria, y por tanto no cabría argumentar que no hay vida humana hasta que esta es única e independiente. Cada vida humana, incluida la de los gemelos monocigóticos quedaría determinada al final de la fecundación, tras la primera división celular como un fenómeno de baja probabilidad derivado del propio proceso de la fecundación. Con toda propiedad se ratificaría que cada vida humana parte de una única célula, bien sea el cigoto o cada una de las células procedentes de su segmentación, que en cualquier caso seguirían un proceso de desarrollo paralelo e independiente desde el principio.
Esta explicación es coherente con el hecho de que una vez constituida la identidad genética estamos ante una nueva vida humana, que podría convertirse en dos o incluso ocasionalmente en más de dos, si por accidente se produjera la gemelación. En cualquier caso, debe sostenerse la identidad genética como el argumento principal para considerar que estamos ante una vida humana.
La indivisibilidad no ha de considerarse requisito para el reconocimiento de la individualidad. El embrión temprano podría ser potencialmente divisible, pero sigue siendo una etapa del ciclo biológico humano, que comenzó tras la fecundación. El embrión, divisible o no, es ya una vida humana que posee su identidad invariable de por vida de la que depende su desarrollo como un ente biológico.
La diferenciación sexual en el hombre
Por su interés en los debates bioéticos relacionados con la distinción entre sexo y género, conviene conocer la base biológica de la diferenciación sexual humana. En los tratados de embriología y de genética humana, se habla de varios criterios de asignación de sexo:
- Sexo cromosómico: ♀ 46, XX y ♂ XY.
- Sexo genético: ♀ sin el gen SRY y ♂ con el gen SRY.
- Sexo gonadal: ♀ con ovarios y ♂ con testículos.
- Sexo psicológico o de crianza.
Las tres primeras denominaciones se corresponden a la realidad biológica de cada ser humano. Sin embargo, el llamado sexo de crianza no está determinado genéticamente, sino que obedece a factores ambientales que influyen en la psicología y el comportamiento en lo referente a lo que se ha dado en llamar orientación sexual, pudiendo diferir del sexo biológico.
Veremos aquí cómo tiene lugar el desarrollo del aparato genital masculino y femenino durante la embriogénesis humana.
La realidad sobre el sexo en el ser humano, es que depende fundamentalmente del desarrollo del aparato genital y de los órganos secundarios masculinos o femeninos, dependientes del programa genético establecido desde la fecundación. La diferenciación sexual está determinada en primer lugar por la constitución cromosómica del llamado par sexual: XX en la mujer o XY en el varón. Esta se establece en la fecundación como se representa en la Figura 3. El sexo heterogamético es el varón, ya que genera dos tipos de espermatozoides, unos con el cromosoma X y otros con el Y, al 50 %, mientras que en la gametogénesis femenina solo se originan gametos de un mismo tipo, todos portadores del cromosoma X. De este modo queda garantizada una descendencia con una equivalente proporción de sexos: 50% varones y 50% mujeres.
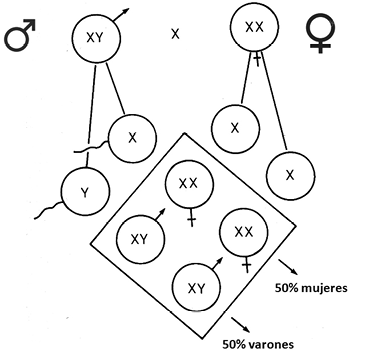
Figura 3. Segregación de los cromosomas sexuales en la gametogénesis masculina y femenina y las combinaciones posibles en la descendencia: 50% varones - 50% mujeres.
El significado biológico de la diferenciación sexual es garantizar la reproducción, para lo que es necesaria la participación de dos sexos con sustanciales diferencias en el aparato productor de los gametos. Pero para garantizar la descendencia, no solamente debe haber diferencias biológicas sino también psicológicas, por los diferentes papeles que han de representar los dos parentales en el cuidado y educación de los hijos. Por ello, en el hombre, el sexo, que se va a desarrollar a partir de la fecundación, se reflejará en el tipo de aparato reproductor masculino o femenino y también en el tipo de cerebro de varón o de mujer102. Ambas cosas suceden de forma genéticamente regulada y en fases cronológicamente distintas del desarrollo embrionario y fetal.
Un embrión humano empieza a desarrollar sus órganos reproductores entre las semanas cuarta y quinta después de la fecundación, con la aparición en el embrión de una gónada primitiva indiferenciada, como vemos en la Figura 4. Después, se complementará con el desarrollo de las gónadas masculina o femenina. Más adelante, se añadirán modificaciones también en el cerebro hasta la vigésima semana del embarazo.
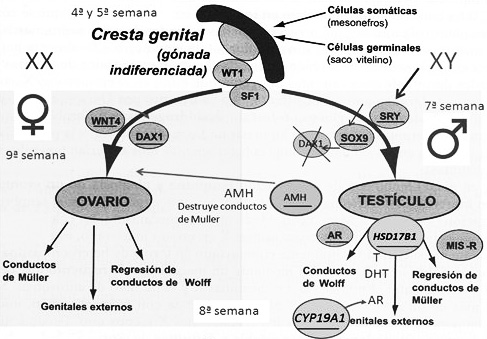
Figura 4. Principales actividades genéticas que determinan la diferenciación gonadal y sexual en el hombre (los símbolos de los genes principales figuran subrayados).
El fenotipo sexual de varón o mujer, empieza en la séptima semana del desarrollo, cuando se expresan los genes reguladores de la gonadogénesis. Hasta ese momento, lo que había es el esbozo de una gónada primitiva potencialmente bisexual, que si no recibe un estímulo genético seguirá su camino hacia el lado femenino. Lo que decanta la formación del aparato genital como de varón o mujer, es la presencia y actividad de un gen regulador, el gen SRY, que se localiza en el cromosoma Y, y por tanto solo tiene lugar en los varones. Si se activa este gen, la gónada hasta entonces indiferenciada, seguirá una ruta de desarrollo testicular. En su defecto, daría lugar a ovarios. Por ello, al gen SRY se le llama también conmutador de sexo.
La expresión del gen SRY es solo el inicio de una cascada de expresiones de genes que irán interviniendo de forma ordenada para añadir progresivamente más elementos a la formación de las gónadas masculinas, y más adelante, a los órganos genitales externos. De entre los múltiples genes que se van activando durante el desarrollo de los órganos genitales, como de varón o de mujer hay uno que es inmediato, el gen SOX9, que bajo el estímulo del gen SRY promueve el camino hacia el desarrollo testicular. En su defecto se seguiría una ruta de desarrollo ovárico.
El desarrollo embrionario funciona realmente como una sinfonía. Cada sexo sigue su propia partitura dependiendo de las notas, los genes, que se expresan o silencian en perfecta armonía, para al final dar lugar a las gónadas y órganos genitales externos de acuerdo con las diferentes instrucciones del programa genético de varón (XY) o mujer (XX).
En presencia del gen SRY se inducirá el desarrollo del mesodermo nefrítico, los conductos de Wolf —en la gónada masculina—. Si no está presente surgirán los conductos de Müller y, más adelante, todas las restantes estructuras gonadales femeninas. Entre otros genes, el regulador WNT4 determina la síntesis de una proteína de señalización que activa a otro gen, el DAX1, antagonista de SRY, lo que cierra el paso al desarrollo de los conductos de Wolf, necesarios para la formación de los testículos a partir de la gónada primitiva y determina la formación de los conductos de Müller, propios de las gónadas femeninas.
En presencia del gen SRY, una vez que se ha inducido la diferenciación de los testículos, comienza la secreción de tres hormonas que intervienen en el desarrollo de los genitales masculinos internos y externos. En las células de Sertoli del órgano testicular en formación, se activa el gen AMH, que codifica la hormona antimulleriana —HAM—, que determina la regresión de los conductos de Muller, produciendo un efecto masculinizante. Al mismo tiempo en las células de Leydig se activan otros dos genes reguladores que desempeñan un papel relevante en la gonadogénesis masculina, el HSD17B1, que sintetiza la testosterona y su derivada la dihidrotestosterona —DHT—, hormonas esteroideas predominantes en el varón, y el AR, que codifica para una proteína que actúa como receptor de los andrógenos —AR—.
La testosterona y su derivada, la DHT, se unen al mismo receptor de andrógenos, pero los dos ligandos producen respuestas diferentes. En presencia de estas hormonas se completa el desarrollo de los órganos genitales masculinos internos: próstata, uretra, glándulas bulbouretrales, vesículas seminales, y los externos: el pene, el epidídimo, y los conductos deferentes. En un momento más avanzado del desarrollo fetal, la testosterona controlará la migración de los testículos desde el abdomen hacia el saco escrotal.
De forma paralela y en ausencia de las hormonas masculinas se desarrollan los órganos genitales femeninos, internos: ovarios, trompa de Falopio y el útero, y los externos: el clítoris, la vagina, los labios, etc.
Entre el segundo y el quinto mes del desarrollo fetal, cuando ya están formados los testículos y los ovarios, la testosterona, mediada por la acción de una enzima codificada por el gen CYP19A1, que se llama aromatasa, también secretada por los testículos, entra en el cerebro donde se convierte en estradiol. Tal como se representa en la Figura 5, este actúa sobre los receptores alfa y beta de los estrógenos —ER—, contribuyendo al desarrollo cerebral masculino. Si el nivel de la testosterona es bajo, como ocurre en el desarrollo gonadal femenino, no se produce la masculinización del cerebro103. Además, una α-fetoproteína protege al cerebro femenino de la masculinización cerebral, impidiendo la entrada de estrógenos en el cerebro104.
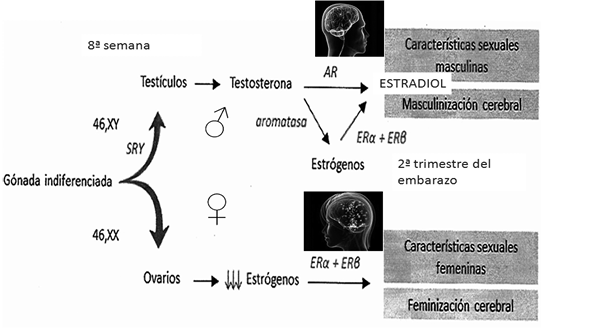
Figura 5. Papel de las hormonas durante la organogénesis gonadal, tanto masculina como femenina (ERα y ERß, son los receptores de estrógenos).
El cerebro es un órgano fundamental en el desarrollo de la sexualidad humana. Ante un elevado nivel de andrógenos durante el período sensible en que han de actuar, las células y tejidos distribuidos por todo el cuerpo deben formar un fenotipo masculino, mientras que si el nivel de andrógenos es bajo se formará un fenotipo femenino.
De este modo, las hormonas influyen en el modelado final tanto de los genitales externos como del sistema nervioso central, determinando los comportamientos psicológicos y sexuales diferentes del varón y la mujer. El hecho de que el desarrollo gonadal tenga lugar en el primer trimestre del desarrollo embrionario-fetal, y el del cerebro, en el segundo trimestre, demuestra la existencia de equipos de genes diferentes pero complementarios al servicio de la reproducción humana105.
Naturalmente el desarrollo de los órganos genitales, condicionado genéticamente, no está exento de la posibilidad de mutaciones en los principales genes que intervienen, o en variaciones de su actividad por factores fisiológicos. Hay casos, como los llamados pseudohermafroditas masculinos, en los que una mutación produce deficiencias en la síntesis de la enzima 5-reductasa, que es necesaria para catalizar la conversión de la testosterona en DHT. A pesar de la secreción hormonal de testosterona, estos varones tienen concentraciones bajas de DHT y, como consecuencia, los genitales externos masculinos y la próstata no se desarrollan completamente durante la vida fetal.
Es evidente que las mutaciones en los genes que determinan la formación de las gónadas, las alteraciones en la constitución cromosómica XX o XY, o la influencia de fármacos, drogas, hormonas u otros agentes, pueden conducir a una alteración en el desarrollo del aparato genital del bebé en fase fetal, dando lugar a los infrecuentes casos de síndromes de malformación de los órganos genitales. Se ha sugerido que las diferencias de sensibilidad a las hormonas sexuales pueden deberse a modificaciones epigenéticas en la región promotora de los genes implicados en la respuesta masculinizante o feminizante, que pueden estar influenciadas por factores fisiológicos internos o causas ambientales durante el desarrollo fetal. En cualquier caso, se trataría de modificaciones epigenéticas no programadas genéticamente.
Usualmente se da por hecho que los genes de un individuo se mantienen invariables a lo largo de la vida, desde la fecundación. Sin embargo, pueden surgir mutaciones genéticas o modificaciones epigenéticas, a veces inapropiadamente denominadas epimutaciones, que pueden determinar un cambio o incluso la anulación total de la expresión de los genes en algún lugar o momento durante el desarrollo embrionario, fetal o incluso en la fase adulta.
Debe tenerse en cuenta que una célula es la unidad funcional mínima de un organismo, y que en un tejido o en un órgano pueden coexistir muchos tipos de células. El hecho es que las diferencias entre ellas son debidas al epigenoma —la parte del genoma que está funcionando—, del que depende el proteoma —las proteínas presentes en la célula—. Ocasionalmente se pueden producir mutaciones puntuales en los genes, o alteraciones cromosómicas numéricas o estructurales que afectan a una célula y sus descendientes. Esto, puede traducirse en la aparición de regiones del cuerpo con diferencias genéticas al resto del organismo, lo que puede manifestarse fenotípicamente dando lugar a un mosaicismo. Los mosaicismos pueden tener consecuencias para la salud y pueden estar implicados en personas con manifestaciones parciales o más o menos acusadas o que afecten a diferentes regiones corporales. Un ejemplo de esto son los falsos positivos o los falsos negativos, para la detección de un síndrome de Down u otras alteraciones cromosómicas, en el análisis de una célula en los embriones durante el llamado diagnóstico genético preimplantatorio o prenatal, de lo que hablaremos más adelante.
También pueden producirse tipos de células alteradas en su función o en su capacidad de proliferación que den origen a la aparición de un tumor. Esto es lo que ocurre en el cáncer, que puede deberse o bien a una mutación en algún protoncogen o en algún gen supresor, que son dos tipos de genes de funciones antagónicas y ambos implicados en la regulación del ritmo de división celular, ya que los protooncogenes tenderían a acelerar el ciclo y los supresores a su moderación. Cualquier alteración en estos genes o en las regiones cromosómicas sensibles que los incluyen, puede determinar la sobreestimulación de la proliferación celular, o la eliminación del freno que la regula, dando lugar a la producción de un tumor.
A los genes, determinantes de los rasgos físicos y fisiológicos de cualquier individuo, se sobreañaden en el ser humano otros factores no genéticos condicionantes de lo que genéricamente se conoce como su personalidad, que es algo que cada individuo se labra tras el nacimiento. Cada ser humano se constituye como consecuencia de una doble contribución que responde a las dimensiones biológica, psicológica y espiritual. Conviene distinguir estas realidades distintas que contribuyen a edificar la vida de cada persona, y especialmente los factores de que dependen. La dimensión biológica se debe a los genes y está determinada por la identidad genética. La dimensión psicológica y la espiritual, se refiere a la conciencia que de sí mismo se forja cada persona y se traduce en una identidad personal, la que de acuerdo con la voluntad e inteligencia más o menos formada, determinará la personalidad y todos los elementos que contribuyen a lo que cada individuo quiere ser, como se muestra en la Figura 6.
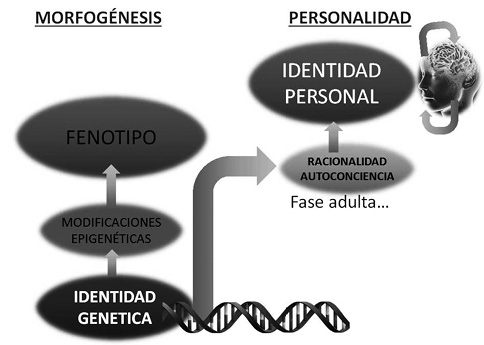
Figura 6. Esquema representando el ascenso en los dos elementos fundamentales de la identidad humana: la identidad genética y la identidad personal.
Tras el nacimiento, durante la autogénesis de la conciencia, crece la percepción individual sobre la realidad que conduce a la construcción de la personalidad. En esta nueva etapa, la percepción de la realidad corporal determinada genéticamente, queda supeditada a la razón y la voluntad de la persona. Al cómo se es, se añade el qué se quiere ser.
Decía el filósofo de la Ilustración David Hume (1711-1766) que la mente humana es una pizarra vacía en la que la experiencia va grabando sus signos. Muchas de nuestras acciones como humanos, y sobre todo las que atañen al comportamiento: bondad, agresividad, instinto abusivo, naturaleza depresiva, orientación sexual, etc. no se heredan, sino que se adquieren. No responden a los dictados del ADN, sino que forman parte de lo que el filósofo español Xavier Zubiri (1898-1983) definía como personalidad, el precipitado que deja en cada persona el contenido de los actos que va ejecutando a lo largo de su vida106. A diferencia de lo que ocurre en el resto de los animales, en cada persona lo que se hace racionalmente es más importante que lo que viene dictado por sus genes.
Hay muchos rasgos de la personalidad que se adquieren a lo largo de la vida, sin duda bajo influencias de carácter ambiental, entendido en sentido amplio como lo que influye en nuestra forma de ser y afrontar la vida. La educación, el estudio, la alimentación, los hábitos de vida, etc., influyen de algún modo en nuestra forma de ser y de pensar, y forjan nuestra conciencia moral y espiritual, pero es nuestra voluntad la que en definitiva está implicada en el modo de afrontar cada realidad. Debido a su carácter no genético, todo lo relacionado con la psicología y la espiritualidad puede variar a lo largo de la vida, dependiendo de las influencias vitales que atraviesa en cada momento cada persona.
Un carácter humano muy dependiente de la personalidad y sobre el que se ha vertido toda clase de opiniones, muchas veces interesadas y apartadas de la realidad, es la inteligencia, que, aunque es difícilmente medible con objetividad, se ha tratado de cuantificar mediante la estimación del llamado coeficiente intelectual —CI—. Se trata de un carácter de heredabilidad media, cuya estimación se basa en ejercicios mentales. Dejando aparte las críticas a los test utilizados para estas estimaciones señalaremos la siguiente consideración llena de sentido común de los genetistas Leslie C. Dunn (1893-1974) y Theodosius Dobzhansky (1900-1975): «Como biólogos, vemos las diferencias humanas como hechos que requieren comprensión e interpretación, no como cualidades que deben condenarse o elogiarse. De hecho, al describir grupos humanos en términos biológicos, no admitimos categorías definidas como bueno o malo, superior o inferior»107.
Está claro que los genes pueden tener que ver con el cómo somos, aunque probablemente menos con lo queremos ser. No entender esto constituyó el gran error que dio origen a las corrientes racistas y eugenésicas de principio del siglo XX y también a las erráticas corrientes actuales de la ideología de género, que ignoran el sexo y lo consideran parte de una herencia cultural en lugar de un hecho biológico del que depende la reproducción.
Todo esto demuestra el interés de la genética en el juicio ético y en particular en el debate bioético. La calificación moral de un rasgo del comportamiento humano puede ser muy distinta de tratarse de una opción voluntariamente aceptada, sin que exista una base genética demostrada, que, de tratarse de un comportamiento inevitable, dependiente de una combinación determinada de genes. Por ello, es necesario salir al paso de aquellos comportamientos humanos que algunos han tratado de justificar bajo la apariencia de su inevitabilidad por razones genéticas. ¿Somos prisioneros de lo que dictan nuestros genes? Es evidente que el ser humano es mucho más que sus genes. La personalidad que se expresa en el modo en cómo se conduce en la vida cada individuo es algo que se construye sobre la base de una experiencia personal.
Las consecuencias de todo lo dicho nos llevan a una reflexión sobre la responsabilidad que los adultos tienen en la educación de los niños. Estos cuando nacen absorben del medio que les rodea la información y las enseñanzas que labrarán su personalidad. En este sentido debe cuidarse especialmente el tipo de información que les llega. Son personas física y psíquicamente vulnerables y su mente, que tardará años en hacerse autónoma para responder libremente a los retos que se les planteen, se configurará conforme a las herramientas intelectuales que durante la infancia hayan adquirido, fruto de las enseñanzas y educación recibida. En este sentido son especialmente el padre y la madre, con sus diferentes papeles complementarios, los únicos responsables de lo que deseen para sus hijos.
De acuerdo con el principio de que el fenotipo es la expresión del genotipo en relación con el medio ambiente, este debe ser especialmente cuidado en las etapas iniciales de la vida y son los padres los responsables de ello.
Otra consideración importante e independiente de lo anterior es que no se debe juzgar a las personas por lo que la naturaleza biológica les haya concedido. Todas las personas son iguales en dignidad con independencia de su edad o condición. Toda persona ha de ser respetada incondicionalmente, con independencia de cualquier característica o aptitud particular que pudiera poseer, simplemente por el mero hecho de ser persona. Nadie es responsable de su identidad genética y por tanto nadie tiene derecho a pensar que un niño o una persona adulta discapacitada, o con una condición mental disminuida, es menos valioso que otro, o que va a ser más o menos feliz en función de una disfunción genética o una patología.
Todas las personas deben considerarse iguales en dignidad y deben tratarse como un fin en sí mismo y respetar su vida, su autonomía y su libertad, haciendo todo lo posible para ayudarles a que se realicen como personas con toda la potencialidad que su constitución biológica les permita. En ningún caso se debe romper el hilomorfismo de la persona, la relación intrínseca entre cuerpo y alma.
Viene al caso señalar especialmente el respeto que se merecen las personas con síndrome de Down, cuya personalidad y desarrollo psicológico devuelve con creces a sus padres, hermanos, familia y a la sociedad que los acoge. Como bien señalaba Jérôme Lejeune (1926-1994), un cromosoma de más o un gen alterado no nos da derecho a menospreciar y aún menos eliminar la vida de un ser humano. Este grave error está en el origen de las corrientes eugenésicas sobre las que trataremos más adelante.