CAPÍTULO 1
ECOSISTEMA PRATENSE
Uno de los objetivos principales que se persigue en el manejo de praderas es mantener una productividad alta. Sin embargo, a menudo, no se conoce en términos cuantitativos la capacidad productiva de la pradera expresada en materia seca u otra unidad similar. Frecuentemente, se pretende conocer la salud ecológica de la pradera independientemente de su producción de tejido vegetal útil expresada en rendimiento de materia seca por unidad de superficie.
Concepto de Condición
La relación entre la productividad potencial de la pradera y la productividad potencial del sitio, se conoce como condición. Esta ha sido definida por Costello (1945) como “el estado de salud de la pradera”, refiriéndose a su salud ecológica. Él aplicó esta definición en la forma más amplia, que incluye tanto la productividad del suelo, como a la vegetación en relación a lo que podría o debería producir bajo condiciones normales de clima y el mejor manejo que se pudiera practicar. El conocimiento de la condición es, según Bailey (1945), esencial para determinar la capacidad sustentadora y planificar el uso de la pradera. El Cuadro 1-1, por ejemplo, muestra varias comunidades uniformadas en las cuales la carga animal es menor cuando se trata de praderas más deterioradas.
La primera mención que se encuentra en literatura pratense sobre el concepto de condición proviene de Sampson, es en los años 1917 y 1919. Según él, el valor de la pradera como productora de tejido vegetal útil, está esencialmente determinado por la etapa de la sucesión. Posteriormente, el concepto de etapa de Sampson fue transformado por condición.
Cuadro 1-1. Capacidad sustentadora de algunas praderas estudiadas por Hutchings y Steward (1953), de acuerdo a su condición
|
Comunidad |
Condición |
|
|
Regular a buena Ha/oveja-mes |
Pobre Ha/oveja-mes |
|
|
Atriplex confertifolia – quenopodiaceae |
1,42 |
2,31 |
|
Atriplex confertifolia – Eurotia lanata – quenopodiaceae |
0,89 |
1,49 |
|
Artemisia nova – Atriplex confertifolia |
0,40 |
0,89 |
|
Eurotia lanata – Chrysothamnus stenophyllus |
0,46 |
1,13 |
|
Cerpocarpus betuloides |
1,13 |
1,74 |
|
Eurotia lanata |
0,40 |
0,97 |
Humphrey (1949), resumiendo las ideas de varios autores y organizaciones, afirmó que, básicamente hay dos puntos de vista para referirse a la condición de la pradera. Uno de ellos es el climático, que se refiere a las condiciones del clima del momento o muy reciente y su influencia en la producción de forraje. Cuando los factores ambientales responsables de la producción de tejido vegetal útil se han presentado en magnitudes favorables para inducir un buen desarrollo de la vegetación, se tiene buena condición de la pradera, o bien, bajo condiciones ambientales, especialmente climáticas, desfavorables, la condición es pobre. Este punto de vista no es de mucho interés para quien esté manejando la pradera.
El otro punto de vista, al cual se refiere Humphrey (1949) es el del potencial de la pradera. Él expresa que la condición de la pradera no es un estado temporal de ella y que, por lo tanto, expresa la producción real de la misma en relación al potencial del lugar. Una pradera en condición buena o excelente produce más tejido vegetal útil, bajo similares condiciones abióticas, que una pradera de condición pobre.
Originalmente, la condición de la pradera para un determinado sitio fue definida por Humphrey (1945 y 1947) como la condición presente de la pradera en relación a la producción que es posible obtener con el mejor manejo práctico. Sin embargo, posteriormente, en 1949, reconoció que aun cuando la clasificación de la condición debe estar basada principalmente en el porcentaje de la producción potencial, hay además otros factores que considerar. La relación que existe entre la potencialidad de producción de forraje y la producción en un determinado momento, fue determinada mediante el uso de indicadores, tales como, la composición botánica, el mantillo y otros.
Condición. Ha sido definida, también, en términos puramente ecológicos. Dyksterhuis (1949) la definió como: el porcentaje de la vegetación presente que es original para el sitio. Después de esto, el concepto ecológico ha sido el más ampliamente usado. Dyksterhuis ha indicado también que, al analizar la condición de la pradera, no debe considerarse el monoclímax como unidad de comparación, aun cuando se consideren períodos muy largos de tiempo. La teoría monoclimácica puede integrar más fielmente todos los componentes del ecosistema y, por lo tanto, representar a un bioma pratense más evolucionado y estable. Sin embargo, Dyksterhuis (1949) ha enfatizado que el concepto de condición tiene una base esencialmente policlimácica, mencionando especialmente el clímax edáfico, fisiográfico y climático. Cuando las praderas se desarrollan en suelos deteriorados o inmaduros, el lento proceso de la génesis del suelo puede hacer que la pradera sea clasificada en condición pobre, regular o buena, en lugar de condición excelente, por un período muy largo de tiempo. Ello se debe a que, a menudo, existen limitantes edáficas o fisiográficas ajenas al manejo mismo de la pradera, las cuales finalmente inducen cambios en los diversos factores bióticos.
La información disponible en este momento hace pensar que la definición de Humphrey es más general e indica realmente lo que es condición. En términos generales, es preferible definir condición en la siguiente forma: es la productividad de tejido vegetal comestible de la pradera en un momento determinado en relación a la productividad potencial del sitio. Condición es, por lo tanto, una proporción entre dos cantidades: una que representa el valor actual de productividad y la otra, el máximo absoluto del sitio. La relación es en base a materia seca producida en ambas etapas sucesionales.
Condición representa una proporción que en sí no es ecológica. Sin embargo, tiene fundamentos ecológicos, porque considera que la productividad potencial y la actual corresponden a dos etapas sucesionales diferentes en una misma sere. Dyksterhuis (1949) lo que hizo en realidad, fue determinar una técnica para clasificar condición basada en la proporción de plantas clímax presente de la pradera, en un momento determinado.
Este punto de vista es parcial y, a pesar de ser un buen indicador para muchas praderas, puede conducir a errores en otras. En verdad, cualquier indicador puede ser utilizado para determinar la condición, ya sean los microorganismos del suelo, insectos y otros grupos de animales inferiores edáficos, microflora, características físicas o químicas del suelo, plantas forrajeras y árboles o arbustos.
Siguiendo las ideas enunciadas por Mayor (1951), se ha concluido que las comunidades vegetales (ν) están relacionadas con cinco factores ambientales:
ν = f (cl, m, r, b, t)
Siendo:
cl: clima regional
m: material generador
r: relieve
b: biota
t: tiempo
El uso de esta ecuación ofrece la posibilidad de buscar relaciones más exactas de los factores determinados en la vegetación y proporcionar los elementos necesarios dentro de los cuales, los datos cuantitativos de la vegetación y demás componentes ambientales pueden ser calculados. Todo esto, fuera de ofrecer símbolos representativos de abstracciones ecológicas, permite, a su vez, la interpretación de la información original.
Cualquier biocenosis puede ser conocida estructuralmente en forma simultánea con su funcionamiento de acuerdo a un solo factor, siempre que los otros no cambien. Así se tiene que, conociendo cualquiera de ellos en un momento determinado, se puede finalmente conocer el valor de ν, por cuanto éste es una consecuencia de los demás. La conveniencia de la aplicación de este principio proviene de las ventajas de utilizar cualquiera de los factores incluidos en la fórmula para la relación de la vegetación potencial.
Todos los elementos del ecosistema cuantitativo del bioma pratense, expresados en la ecuación anterior conducen, finalmente, a la expresión energética del ecosistema. La pradera con su población de ganado y vida silvestre constituye un complejo de comunidades bióticas (Osborn, 1956). La condición de esta biota depende del continuo flujo de energía y sustancias desde el medio inorgánico, a través de los diferentes niveles de vida y hacia el comienzo del ciclo. La magnitud de este flujo depende de la base climática y edáfica de cada asociación ecológica. Generalmente, varía de grande a pequeño, a medida que el clima cambia de húmedo a seco y de cálido a frío.
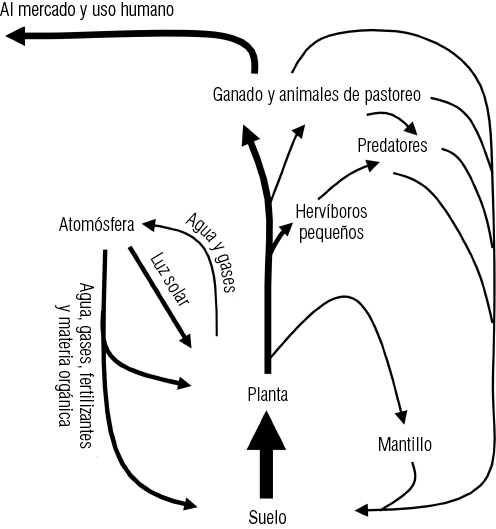
El complemento del ciclo de energía y nutrientes proviene del desprendimiento de elementos en el sitio de la descomposición de restos de animales y vegetales, del intercambio de gases y humedad en la atmósfera e incorporación de ellos al ciclo local y de la captación de energía solar y elaboración de nuevos materiales orgánicos por fotosíntesis (Osborn, 1956). El suministro de energía y nutrientes al ciclo, puede ser incrementado localmente, adicionándole fertilizantes minerales o materia orgánica al suelo, o por la adición de alimentos para el ganado, producido en otros lugares. La continuidad del funcionamiento al máximo de la biota pratense depende, por lo tanto, de tres requisitos fundamentales: el suelo debe ser mantenido intacto, sin pérdidas por erosión u otras causas; el uso continuado de agua y gases de la atmósfera, y el volumen y calidad del tejido fotosintético de la vegetación debe ser mantenido para utilizar la luz solar. La vegetación es el medio que integra todos los componentes que incluye la planta, el suelo y el mantillo (Figura 1-1).
Figura 1-1. Ciclo de energía del bioma pratense (según Osborn, 1956)
La unidad ecológica básica es el ecosistema
El concepto de ecosistema involucra la existencia de una estructura integrada por elementos fundamentales, los cuales están siempre presentes y de un funcionamiento ordenado de esta unidad. La acción que se ejerce sobre el ecosistema, por períodos cortos o largos de tiempo, se traduce en una reacción que termina por modificar la estructura y funcionamiento. Ello es la dinámica del ecosistema.
La estructura ecosistémica consta de dos partes fundamentales: una de naturaleza abiótica o de la materia no viva y otra de naturaleza biótica, en la cual intervienen organismos animales y vegetales en todos sus niveles de integración (Figura 1-2).
El componente abiótico consta de dos partes: los recursos del medio y el hábitat. Los recursos del medio están representados por los nutrientes inorgánicos que el organismo necesita para su subsistencia. Entre ellos se encuentran el carbono, hidrógeno y oxígeno que obtienen del agua y anhídrido carbónico. Ellos, junto con el nitrógeno, fósforo y azufre, constituyen las proteínas con lo cual componen el protoplasma. Además de ellos son esenciales para el crecimiento y desarrollo vegetal: calcio, magnesio, potasio, hierro, manganeso, molibdeno, cobre, boro, zinc, cloro, cobalto, vanadio y sílice.
El funcionamiento del ecosistema requiere, también, del aporte de energía luminosa. El agua es un recurso fundamental del ecosistema. Intervienen en la formación de carbohidratos, aminoácidos, proteínas y protoplasma. En la primera etapa se logra la formación de carbohidratos a través del proceso de fotosíntesis, al combinar agua, anhídrido carbónico y energía luminosa.
6CO2 + 6H2O + 673 calorías  C6H12O6 + 6O2
C6H12O6 + 6O2
En las etapas siguientes otros elementos se incorporan y logran constituir unidades más complejas.
El déficit o exceso de cualquiera de los elementos descritos hace que los organismos alteren su funcionamiento.
El agua interviene en el ecosistema como un recurso fundamental. Su importancia reside, además del aporte de hidrógeno y oxígeno, en otras funciones vitales. Mantiene la turgencia, transporte de sustancias y permite reacciones químicas dentro del organismo.
En manejo de praderas, uno de los mecanismos más utilizados para mejorar la productividad es modificar la tasa de suministro de recursos hasta hacerlo que se aproxime al óptimo. A través del manejo se puede reducir las pérdidas de algunos recursos esenciales del medio, aumentar la circulación y recirculación, hacer más eficiente su utilización a través del paso por los ecosistemas y, por lo tanto, conducirlo finalmente al incremento de la productividad.
Figura 1-2. Arquitectura y funcionamiento del ecosistema (Gastó, 1979)
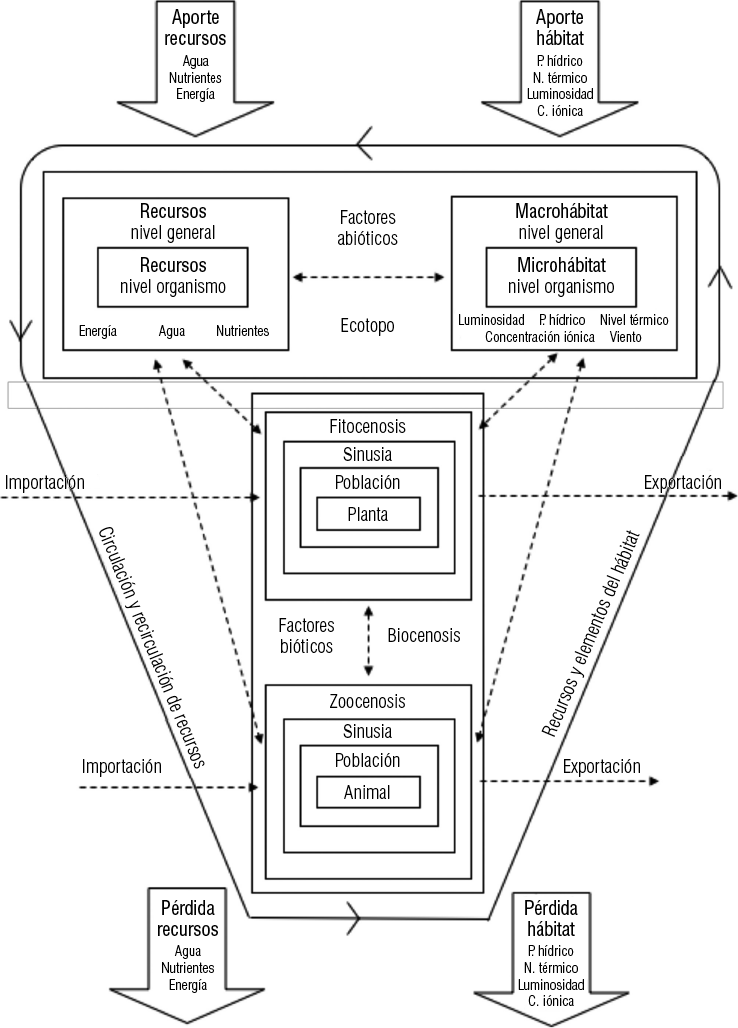
Los nutrientes minerales circulan en una alta proporción. Algunos de ellos pasan por el ecosistema y no logran recircular, ya sea debido a su naturaleza propia o al mal manejo de la unidad pratense. El proceso de circulación y recirculación de elementos a través del medio biótico y abiótico recibe el nombre de ciclo biogeoquímico. Un porcentaje pequeño de los elementos se pierde en forma natural fuera del ecosistema a través del aire, del agua o son fijados en las capas edáficas más profundas.
Algunos recursos no circulan en el ecosistema, sino que pasan a través de él. En esta categoría se incluye la mayor proporción del agua, energía solar y de algunos elementos tales como el carbono, oxígeno y nitrógeno. Sólo una proporción de estos recursos, que puede ser muy baja, recircula en el ecosistema.
El aporte de recursos desde fuera del ecosistema es de máxima importancia. La cantidad aportada puede ser el elemento limitante de la productividad. Aquellos sistemas que se encuentran en equilibrio tienen un aporte de recursos desde fuera del ecosistema igual a la pérdida.
Aporte de recursos – Pérdida de recursos = 0
Los ecosistemas que se encuentran en alguna etapa seral distinta de cualquier tipo de clímax tienen valores diferentes a cero y, por lo tanto, no están en equilibrio. Si el valor es mayor o menor de cero es indicativo de que existe un proceso de sucesiones progresivas o retrogresivas.
Aporte de recursos – Pérdida de recursos ≠ 0
En un ecosistema pratense, manejado antropogénicamente, se persigue comúnmente como finalidad la de exportar fuera del sistema, productos vegetales o animales, para ser utilizados por el hombre en otros lugares. Además, a menudo se recurre a importaciones de biomasa de la misma naturaleza que se introduce temporalmente en el sistema por un período hasta que logra alcanzar mayor desarrollo. Los ecosistemas pratenses exportan comúnmente ganado de cierto desarrollo, o incluso, puede exportar pasto verde o heno. Algunos ecosistemas importan ganado de menor desarrollo o peso para luego exportarlo algún tiempo después. Para que exista equilibrio en un ecosistema pratense, los aportes de recursos más las importaciones deben ser iguales a las pérdidas de recursos más las exportaciones.
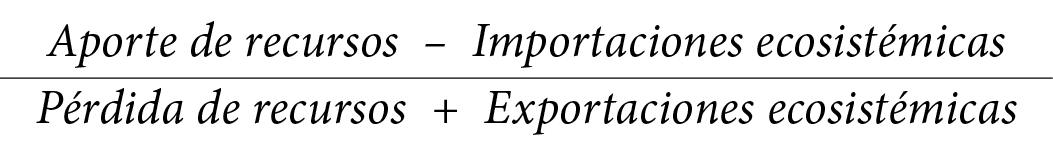 = 1
= 1
Frecuentemente, no es posible suplementar antropogénicamente el suministro de recursos al ecosistema natural. El mejoramiento del manejo ecosistémico, permite a menudo mejorar la eficiencia de utilización de los recursos, disminuir las pérdidas e incrementar la productividad y capacidad de exportación de productos pratenses.
La cuantificación del aporte y pérdidas de recursos puede ser una forma de evaluación de praderas, siempre que se mantengan constantes otros factores. La productividad del ecosistema se incrementa al aproximarse al óptimo la relación entre el suministro, las pérdidas y las necesidades de recursos.
Continuando con el esquema presentado en la Figura 1-2, se presenta el microhábitat, como el lugar donde vive el organismo. La cuantificación y descripción del medio donde vive un organismo interesa en el lugar que le rodea en forma inmediata.
El macrohábitat es el lugar general donde se desarrolla el organismo. Depende del macroclima y del sustrato original del lugar, sin considerar la acción recíproca del organismo sobre el medio y del medio sobre el ser vivo. El resultado de la acción, reacción y coacción en el desarrollo del microhábitat, a menudo es muy diferente del macrohábitat general de la región. Los organismos se adaptan al microhábitat donde viven, independientemente del macrohábitat general de la zona, el que a menudo sobrepasa sus límites de tolerancia.
La alteración del ecosistema puede llegar a niveles tales que concluya por modificar el microhábitat particular, hasta niveles tales que sus características se aproximen en exceso o defecto al macrohábitat y lo hagan así intolerable para el organismo, la población, la sinusia o la biocenosis original del lugar. Se tiene en esta forma, que organismos adaptados a ecosistemas bien manejados, desaparecen al ser mal manejados. Ello se interpreta como una inadaptación al hábitat, ignorándose su capacidad adaptativa a microhábitats favorables.
El microhábitat está definido principalmente por la intensidad luminosa en la cual el organismo vive y se desarrolla, el potencial hídrico existente en el medio circundante al organismo y la concentración iónica. Además, el nivel térmico y la velocidad del viento son elementos que describen las características ambientales del lugar. El organismo no utiliza como recurso a ninguno de ellos, pero sus límites de tolerancia ambiental no le permiten vivir en un hábitat determinado cuando el nivel de cualquiera de ellos es demasiado elevado o bajo. Por no constituir una parte integrante del organismo no se les considera como recursos, sino como hábitat.
El medio abiótico original de un sistema puede ser invadido por organismos vegetales. El establecimiento de algún ejemplar aislado no se considera exitoso, mientras la especie no logra formar una población. La población es un grupo de organismos de la misma especie, limitados en espacio y tiempo. La población es diferente del organismo, por cuanto tiene tasa de natalidad, mortalidad y crecimiento, y el individuo no los tiene. Los grupos de poblaciones de la misma forma vital constituyen una sinusia y su conjunto es la fitocenosis en el caso de los vegetales y la zoocenosis en el caso de los animales. Ambos constituyen la biocenosis.
La comunidad vegetal o fitocenosis tiene en cada caso una estructura individual, poblacional y sinusial característica. Ellos son el resultado de un proceso de adaptación y adecuación al ecosistema en su conjunto, donde se integran los recursos, el hábitat, la fitocenosis y la zoocenosis. La acción directa o indirecta del usuario de este recurso, el hombre, modifica al ecosistema y con ello altera su productividad y estabilidad.
La zoocenosis depende de la fitocenosis, de donde deriva su alimento. La acción de la fauna sobre los vegetales altera considerablemente su funcionamiento, lo que puede conducir a un mejoramiento o deterioro de la biocenosis en su conjunto. Los niveles de integración de la zoocenosis son equivalentes a los de los vegetales. La alteración de la acción de la zoocenosis sobre la vegetación es, a menudo, el elemento más importante que regula la estructura, el funcionamiento y la dinámica del ecosistema pratense.
La alteración de cualquiera de los cuatro elementos principales del ecosistema modifica directa o indirectamente a todos los demás y, por consiguiente, conduce a un clímax diferente. El esquema de tales cambios y relaciones está presentado en la Figura 1-2. Las flechas indican sus relaciones. El ecosistema se modifica holocenóticamente.
La pradera es un complejo de partes íntimamente relacionadas y que normalmente pueden estar en equilibrio entre ellas (Ellison, 1949). Como resultado de estas interrelaciones y, debido a las reacciones de las plantas y animales con el medio, ocurren cambios ordenados sin la pérdida de la integridad del complejo o sucesiones primarias. Estos cambios son demasiado lentos como para ser manipulados en forma práctica con el manejo de la pradera.
Las sucesiones secundarias o subseres resultan como consecuencia de la perturbación del complejo. Son más rápidas que las priseres o sucesiones primarias y pueden ser manipuladas al manejar la pradera y producir resultados deseables en un período razonable de tiempo (Ellison, 1949). Cuando los cambios son tan extremos como para destruir la integridad del complejo por erosión acelerada u otras causas, el término sucesión ya no es apropiado y se produce lo que se conocce como cambios catastróficos.
La utilización del concepto de condición significa la aceptación a priori de algunas premisas que, generalmente, no están explícitamente descritas o no son universalmente aceptadas. La primera de ellas se relaciona con la identidad de la composición botánica. En este sentido, se utiliza generalmente la especie como unidad, pero es conveniente sobrepasar los límites convencionales de clasificación taxonómica y utilizar ecotipo, ecoclino o biotipo que significa generalmente, similaridad genética dentro de la población.
El mecanismo de evolución de las taxas aparece esquemáticamente representado en la Figura 1-2. En ella se observa que las sucesiones vegetales desempeñan su rol modificando el hábitat y nicho a medida que las progresiones se desarrollan.
La estructura de la comunidad se ve cada vez más compleja a medida que el nicho de la taxa se hace más limitado, pero, al mismo tiempo, más específico. Paralelamente a ello, el hábitat y el microhábitat son modificados, creando condiciones bióticas y abióticas que difieren de una etapa sucesional a la siguiente.
Este medio, es el mecanismo seleccionador de poblaciones, tanto vegetales como animales. La variabilidad interna proviene de modificaciones genéticas de los individuos, ya sea causada por recombinación de genes, mutaciones o cualquier otra causa. Los individuos que se formen en la población obedecen a una constitución genética bien determinada y son capaces de reaccionar morfológicamente a estímulos del medio biótico y abiótico circundante. Se materializa así, un cambio en la composición genética de la población natural, la que finalmente, se presenta con características diferentes a las de la población original. Esta nueva población, a su vez, induce modificaciones ulteriores a las sucesiones vegetales hasta que, por último, se produce una relativa estabilidad sucesional y evolutiva.
Si se enfoca el aspecto de las taxas desde un punto de vista policlimáxico o de continuum, es posible pensar que aquellas comunidades que corresponden a diferentes policlímax están constituidas, también, por taxas muy variadas, aun cuando a menudo incluyen las mismas especies. Esto es lo que se desprende de los trabajos de McMillan, (1960) y Workman y West (1967 y 1969).
La gradiente genética de muchas especies es la clave de la continuidad de un tipo vegetacional sobre la gradiente hábitat y el nicho. Los ecotipos y ecoclinos de una comunidad permiten el ajuste vegetacional al ecosistema donde se desarrollan, como asimismo a las condiciones de manejo a que son sometidos. Por lo tanto, aun en una comunidad pratense estabilizada pueden estar ocurriendo evoluciones sucesionales en la población de algunas especies dominantes hasta alcanzar la etapa de equilibrio, tanto evolutivo como sucesional.
La información bibliográfica acumulada permite concluir que, la remoción del follaje resulta generalmente un daño para la población (Ellison, 1960). De allí que el efecto dañino diferencial sobre las diversas poblaciones y ecotipos que constituyen la comunidad pratense se traduce, finalmente, en un ajuste diferente de los componentes, debido a la interacción y consortismos.
Los resultados presentados por Peterson (1962) confirman la evidencia, que algunos cambios que se producen en la pradera bajo condiciones de pastoreo pesado prolongado, favorecen la persistencia de algunas gramíneas, por razones de índole variada: crecimiento postpacimiento relativamente rápido, de rebrote de yemas más vigorosos y verdes, mantenimiento de reservas alimentarias o de carbohidratos en la planta moderadamente altas a muy altas, crecimiento durante la iniciación de la estación más bien retardado, pudiendo así escapar del pastoreo excesivo y, finalmente, un crecimiento más postrado o rastrero.
Las estructuras genotípicas de la población son afectadas por las diferentes intensidades y frecuencias de pastoreo. Las características morfológicas de una población de ballica de rotación corta (Lolium perenne x Lolium multiflorum), demostraron ser diferentes de acuerdo al sistema de pastoreo al cual la población había sido previamente sometida. Bajo condiciones de pastoreo frecuente e intensivo aumentó la proporción en la población de individuos de morfología similar a Lolium perenne, pero bajo pastoreo liviano y distanciado se incrementó la preponderancia del tipo Lolium multiflorum (Brougham, 1960).
Algunos ecotipos de Dactylis glomerata presentan mayor persistencia y sobreviven pastoreos más intensivos que otras poblaciones de la misma especie. En un estudio conducido por Kemp (1937), se encontró que todos los ecotipos resistentes al pastoreo intensivo tenían solo una característica en común, que todas las yemas, sin importar su ubicación en las plantas, tienden a crecer horizontalmente antes de girar hacia arriba para permitir el desarrollo de un culmo floral erecto. Este desarrollo procumbente de los culmos es el responsable de la resistencia al pacimiento del ganado, lo cual permite que ellos utilicen solo las hojas del crecimiento basal. Los ecotipos menos resistentes al pastoreo presentan crecimiento inicial erecto de sus yemas y elongación temprana de los culmos florales.
Es conveniente distinguir entre las etapas sucesionales en equilibrio, tales como policlímax, disclímax, paraclímax o simples, etapas sucesionales que, a menudo, pueden presentar una composición botánica similares, pero que realmente corresponden a seres diferentes. En el caso de las etapas sucesionales en equilibrio, el retardo o histéresis es igual a cero, o bien, a veces se puede comparar con magnitudes similares de retardo. Lo dicho anteriormente acerca del retardo no rige cuando se trata de comparar etapas sucesionales diferentes dentro de una misma sere. Bajo tales circunstancias se compara la etapa sucesional correspondiente con un clímax que se sabe en equilibrio y cuyo retardo es igual a cero, con una etapa sucesional cualquiera, cuyo retardo es desconocido, pero cuya correlación entre el o los factores vegetacionales indicadores (condición) y la productividad real están perfectamente correlacionados y la curva de regresión determinada, de acuerdo a métodos estadísticos universalmente aceptados.
No parece conveniente simplificar la clasificación de la condición en solo cuatro categorías: excelente, buena, regular y pobre. Es preferible utilizar una relación cuantitativa entre condición y productividad. Cualquier valor de la composición botánica o condición, en general, puede ser determinado como un valor que no corresponde necesariamente a una de las cuatro clases, sino que representa una magnitud mensurable numéricamente. Por razones prácticas, en algunos casos, puede calcularse solamente la categoría de la condición de la pradera, pero en teoría debe determinarse primeramente su magnitud exacta.
No es conveniente, utilizar siempre como único fitómetro de comparación al clímax. En algunos casos, su utilización no es conveniente ni práctica, pero en otros, por definición es totalmente imposible. Tal es el caso de las praderas sucesionales en general, cuyo clímax, a menudo, no es pradera y, por lo tanto, toda comparación sería imposible. En este caso, las comparaciones deben hacerse en etapas sucesionales bien conocidas y muchas veces en función del tiempo. Bajo circunstancias tales como las presentadas en esta sección, la velocidad de la sucesión puede introducir una dimensión de valor en la evaluación de la condición. Este grupo de comparaciones es especialmente valioso en todos aquellos casos en que se analizan praderas culti-sucesionales. Se ha demostrado que las progresiones y retrogresiones sucesionales no ocurren indistintamente a la misma velocidad y duración. Los cambios que se producen en la vegetación pueden proceder en diversas direcciones dependiendo del carácter de la presión de pastoreo aplicada (Ellison, 1960).
El concepto de condición es realmente mucho más amplio que el significado que se le ha dado en la literatura pratense. En ecología general ha sido descrito y usado implícitamente en forma frecuente, pero no basta con ello y es necesario, en este momento, una mayor difusión de su alcance en otras ramas de la biología aplicada. Puede aplicarse a cualquier tipo de praderas o biomas productores de tejido vegetal útil, de aprovechamiento directo o indirecto por el hombre, o indirectamente por herbívoros de utilización humana. La definición del concepto en forma teórica y abstracta es de gran importancia para comprender la real magnitud de sus aplicaciones, como la capacitación para la utilización en un sentido más amplio.
El patrón de comparación de la productividad potencial del sitio no tiene por qué ser exclusivamente el clímax en el sentido policlimáxico, generalmente aceptado. Debe ser la etapa sucesional que en forma sostenida sea capaz de producir la mayor cantidad de tejido vegetal pratense utilizable. Esto puede ocurrir, ya sea en el ambiente natural o modificado parcial o totalmente. En términos económicos, la comparación debe hacerse con la etapa que, en forma sostenida sea capaz de producir la mayor cantidad de utilidad expresada en unidades monetarias. Si así ocurre, el ideal de condición puede variar de acuerdo al desarrollo de las ciencias agropecuarias y forestales o de la situación económica y social del país.
Tradicionalmente la composición botánica ha sido el indicador de condición más frecuentemente usado. Sin embargo, puede usarse también como indicador de condición, otros elementos tales como relaciones hídricas, mantillo, etc. Si se trata de utilizar las relaciones hídricas, la etapa óptima o patrón de comparación es aquella que, en forma sostenida sea capaz de producir una alta cosecha y calidad de agua.
Condición, entonces, debe ser definida como el porcentaje de la composición botánica y precisamente la especie predominante característica de la etapa sucesional óptima.
Mucho se ha mencionado en la literatura pratense de los cambios que se producen en los factores ambientales y bióticos a medida que se producen sucesiones progresivas y retrogresivas. Sin embargo, los estudios ecológicos relacionados con praderas, no son muy claros en este sentido. Varios autores (Odum, 1963; Cooke, 1967 y Mergalef, 1958), han resumido en forma de un modelo tabular los cambios que se producen en el desarrollo sucesional. Algunos de los atributos del ecosistema en las etapas sucesionales prístinas o iniciales y en las maduras, cuando el ecosistema se aproxima al clímax, aparecen presentados en el Cuadro 1-2.
Cuadro 1-2. Tendencias que se presentan en el desarrollo sucesional de ecosistema según datos presentados por Odum (1963), Mergalef (1958) y Cooke (1967). La columna Condición es un agregado del autor y solo representa su interpretación en relación a lo que implícita o explícitamente propone la literatura pratense
|
Atributos del Ecosistema |
Sucesiones en general |
Condición |
||
|
Iniciales |
Maduras |
Pobre |
Excelente |
|
|
Energía de la comunidad |
||||
|
> 1 (autótrofas) < 1 (heterótrofas) |
1 aproximadamente |
Baja |
Alta |
|
Alta |
Baja |
Varías según sere y etapa óptima |
|
|
Alta |
Baja |
Baja |
Alta |
|
Lineal (predominantemente pastoreo) |
Reticular (predominantemente detritus) |
Lineal (predominantemente pastoreo) |
Lineal (predominantemente pastoreo) |
|
Alta |
Baja |
— |
— |
|
Alta |
Baja |
Baja |
Alta |
|
Alta |
Baja |
— |
— |
|
Baja |
Alta |
— |
— |
|
Alta |
Baja |
— |
— |
|
Baja |
Alta |
— |
— |
|
Estructura de la comunidad |
||||
|
1. Biomasa en Pie y Materia orgánica |
Baja |
Alta |
— |
— |
|
2. Diversidad de Especies |
Baja |
Alta |
Baja |
Alta |
|
3. Diversidad Bioquímica |
Baja |
Alta |
— |
— |
|
4. Estratificación |
Poco desarrollada |
Bien desarrollada |
Poco desarrollada |
Bien desarrollada |
|
Historia Vital |
||||
|
1. Especialización del Nicho |
Amplia |
Ajustada |
Amplia |
Ajustada |
|
2. Tamaño de organismos |
Pequeños |
Grandes |
— |
— |
|
3. Ciclo de vida |
Cortos |
Largos |
Cortos |
Largos |
|
Ciclo de nutrientes |
||||
|
1. Nutrientes inorgánicos |
Alta |
Baja |
Alta |
Baja |
|
2. Ciclos minerales |
Abierto |
Cerrado |
Abierto |
Cerrado |
|
3. Tasa de intercambio de nutrientes. Organismo-medio ambiente |
Rápido |
Lento |
Lento |
Rápido |
|
4. Papel del animal en la regeneración de nutrientes |
No importante |
Importante |
No importante |
Importante |
|
Eficiencia |
||||
|
1. Mantención del sistema |
Baja |
Alta |
Baja |
Alta |
|
2. Producción |
Alta |
Baja |
Baja |
Alta |
|
3. Clorofila (N° de asimilación) |
Baja |
Alta |
— |
— |
|
Homeostasis Total |
||||
|
1. Simbiosis interna (interdependencia de organismo) |
Baja |
Alta |
Baja |
Alta |
|
2. Conservación de nutrientes |
Mala |
Buena |
Mala |
Buena |
|
3. Estabilidad (resistencia a la perturbación externa) |
Mala |
Buena |
Mala |
Buena |
|
4. Entropía |
Alta |
Baja |
— |
— |
|
5. Información |
Baja |
Alta |
— |
— |
|
Estabilidad |
||||
|
1. Metabolismo |
Baja |
Alta |
— |
— |
El modelo tabular de sucesiones presentado, indica categóricamente que no puede existir una etapa clímax en la cual la proporción entre producción bruta y respiración de la comunidad sea distinta de 1. En sucesiones heterótrofas, donde la respiración excede por un margen amplio a la producción bruta, esta proporción es muy pequeña en las etapas iniciales, pero aumenta a medida que la sere madura, cuando alcanza un valor cercano a 1.
En las sucesiones autótrofas, que es el caso de la mayoría de las praderas, ocurre sin embargo lo inverso. El valor de la proporción es en un comienzo muy alto, mucho mayor que 1, ya que debe producirse un exceso de materia orgánica y de biomasa en pie de manera de modificar el medio ambiente. Estas modificaciones graduales que ocurren en un ecosistema desbalanceado son, en realidad, el motor impulsor del avance sucesional.
No puede existir sucesión en un ecosistema natural, a no ser que se produzcan modificaciones autógenas en él. De todos los parámetros que pueden determinarse en una sucesión, la proporción Pb/R es, probablemente la más significativa. Ella evalúa en forma integral la totalidad de la energía, a través de la producción bruta del ecosistema, en un momento determinado, y los factores subtractivos o degradantes de la energía que se representan por la respiración total del ecosistema. El consumo total de oxígeno es un índice de la productividad bruta del ecosistema. En cuanto a la pionera, tiene el menor consumo de los tres. La vegetación clímax, sin embargo, por su naturaleza misma, no puede producir modificaciones autógenas y la mayor producción bruta tiene que ir acompañada de un mayor consumo de material energético en las funciones vitales, lo que se manifiesta por un mayor flujo de energía a través del cauce detritófago (Figura 1-3).
Figura 1-3. Consumo total de oxígeno por microorganismos del suelo, en tres etapas sucesionales diferentes y en diversas épocas del año (según Mallik y Rice, 1966)
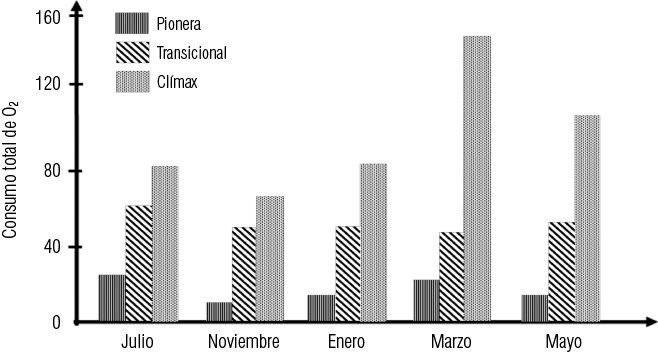
De lo discutido en los párrafos anteriores se desprende que, en la etapa clímax, producción bruta menos respiración tiene que ser igual a cero, pues de lo contrario se estarían produciendo modificaciones autógenas en el ecosistema global. No puede haber modificación global en el ecosistema y haber, al mismo tiempo, equilibrio. Ahora bien:
Producción bruta del ecosistema – Respiración del ecosistema = Producción neta del ecosistema
Si se tiene una sere en la etapa sucesional clímax
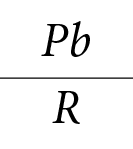 = 1
= 1
por lo tanto, Pb = R
por lo tanto, Pb – R = 0
Si Pb – R = Pn
Pn = 0
Esto es lo que ocurre en el clímax. No puede pretenderse entonces, mantener una pradera en etapa clímax, por cuanto la producción neta de ella sería igual a cero y eso es, precisamente, lo que no se persigue.
Condición, en la forma tradicionalmente definida es la proporción entre una pradera cualquiera, en un momento determinado, y el clímax respectivo, siendo la pradera clímax lo óptimo que puede esperarse. Esto no puede ser así. Lo que en realidad se persigue en manejo de praderas es una etapa sucesional distinta del clímax, en la cual exista estabilidad del medio abiótico o al menos no se presenten cambios destructivos o retrogradantes, en la que se obtiene un máximo valor de la relación Pb/R y de Pb–R, expresado en tejido vegetal útil.
Se presenta, por lo tanto, una controversia. Si el clímax es pradera y este se alcanza sin pastoreo de mamíferos domésticos superiores, un mejoramiento de la condición significa un aumento de las especies clímax simultáneamente con la magnificación de los organismos detritófagos. Según Harlan (1959), las especies clímax son menos susceptibles de pastoreo y, por lo tanto, conducen a una composición botánica menos favorable. Según este autor existen diferencias fundamentales entre las especies pratenses, clímax y disclímax y ello es una característica de la especie. Las especies clímax se comportan como tales en cualquier lugar que crezcan. Así se tiene que, las especies pratenses más importantes en el mundo son disclímax. Los individuos de este grupo de especies se desarrollan bien bajo condiciones alteradas y soportan el pastoreo y corte. Son buenas competidoras con otras especies cuando se utiliza la pradera, pero tienden a desaparecer cuando se les mantiene en exclusiones. Las especies pratenses clímax no se comportan bien en condiciones alteradas y tienden a desaparecer bajo condiciones de pastoreo y corte, responden modestamente a niveles altos de fertilizantes y son productoras pobres y erráticas de semillas. Probablemente, la mayor parte de las especies clímax han evolucionado en ausencia de mamíferos herbívoros, los cuales, a su vez, no han podido evolucionar en estas condiciones, debido a que destruirían o afectarían severamente la vegetación (Harlan, 1959).
Las condiciones ambientales y de manejo pueden ser responsables de la persistencia de las poblaciones que componen la biocenosis pratense. Las especies pratenses están supeditadas a las condiciones ambientales, las cuales determinan, en última instancia, si la reproducción de los individuos se realiza sexual o vegetativamente. La ubicación del meristema apical unido al largo del día son, a menudo, responsables del tipo de reproducción que presenta (Morgan y Grover, 1940).
No existen motivos justificados para promover sucesiones progresivas hacia el clímax, ya que las especies disclímax son más susceptibles de utilización por herbívoros. Por lo tanto, el avance sucesional en las etapas próximas al clímax significa proveer organismos vegetales más susceptibles y, probablemente, menos productivos. En praderas naturales, el principal objetivo es mantener un disclímax, pero en la etapa de mayor producción de tejido vegetal útil.
La cadena alimentaria que existe en la mayoría de los ecosistemas prístinos es de naturaleza lineal, en la cual, el tejido vegetal es utilizado principalmente por herbívoros a través del pastoreo. En etapas sucesionales más avanzadas, a medida que se produce un acercamiento al clímax, la cadena se transforma de lineal a reticular y, de predominantemente de pastoreo en predominantemente de detritus. La condición buena o excelente no puede significar un incremento de la utilización de detritus en desmedro del pastoreo. Al contrario, cualquier pradera en condición buena o excelente, debe representar el máximo de pastoreo y el mínimo de detritus. La producción de detritus debe ser solo lo necesario para mantener una buena conservación de los recursos edáficos y demás elementos abióticos.
El avance sucesional de la comunidad conduce, necesariamente, a una mayor complejidad de los componentes vegetales. Estos significan mayor eficiencia, por cuanto los medios disponibles son más íntegramente utilizados si existe una mayor variabilidad de especies, lo que se logra en etapas cercanas al clímax. Dwyer (1958) encontró que ninguna de las hierbas pivotantes que existían en la pradera de gramíneas que estudió, significaban disminución del rendimiento de la especie angustifoliada. La latifoliada consumía el agua y los nutrimentos debajo de la estrata radicular correspondiente a la gramínea y, en esta forma, reduciendo la competencia edáfica al mínimo.
La fotosíntesis neta de la comunidad es alta en ecosistemas prístinos y baja en ecosistemas maduros, ya que las modificaciones autógenas son mínimas. Sin embargo, si este fuera el caso en praderas clímax o en condición excelente, significaría una reducción de la eficiencia del ecosistema y ello está divorciado de los objetivos perseguidos con un buen manejo de praderas. La producción neta de tejido vegetal útil debe ser máxima en praderas en condición excelente.
La tasa de intercambio de nutrientes disminuye a medida que se produce un acercamiento sucesional hacia el clímax. Es lógico que así sea, pero en manejo de praderas se pretende utilizar el ecosistema en forma tal de obtener una rápida tasa de intercambio de nutrientes con el objeto de incrementar su eficiencia y maximizar la producción de tejido vegetal útil. No puede, por lo tanto, pretenderse manejar una pradera de etapas sucesionales clímax cuando se sabe que eso significa, al mismo tiempo, una reducción de la tasa de intercambio de nutrientes. El buen manejo de praderas persigue una rápida tasa de intercambio de nutrientes en el ecosistema.
La producción neta de ecosistemas en etapas sucesionales maduras, es baja y disminuye a medida que se acerca hacia el clímax. En las etapas sucesionales prístinas, en cambio, la producción neta es alta. Nuevamente aquí se tiene una contradicción, ya que buen manejo significa alta producción y, por lo tanto, no puede pretenderse mantener la pradera clímax en forma excelente, pues su producción neta tiene que ser baja. El objetivo principal en el manejo de praderas es alta producción.
Considerando en conjunto el modelo tabular presentado anteriormente en la literatura pratense considerada en este trabajo, se puede concluir que, lo que se persigue con un buen manejo de praderas no es una utilización que conduzca hacia una etapa estable, final y única, el clímax. Condición excelente se produce, generalmente, en etapas sucesionales avanzadas o maduras, pero nunca en el clímax. Avance sucesional significa inicialmente, mejoramiento de la condición hasta que se alcanza un grado de desarrollo óptimo pratense.
Progresiones superiores al óptimo o condición excelente significan un deterioro pratense, aun cuando en un sentido ecológico estricto pueda significar un avance o mejora.
Clasificación sucesional de las praderas
A continuación, se presenta un resumen de esta clasificación.
Praderas Sucesionales
• Priseres
1. Naturales
– Praderas de herbáceas perennes en clímax bosque
· Hidrosere
· Xerosere
– Pradera anual en clímax matorral
– Pradera anual en clímax herbáceo perenne
– Pradera anual en clímax bosque
– Psamosere pratense
2. Inducidas
– Psamosere pratense
• Subseres
1. Cultisucesiones pioneras
– Con aporte adicional de disemínulas
· Temporales
· Anuales y bianuales
· Rotación corta
· Rotación larga
– Sin aporte adicional de disemínulas
· Praderas de malezas o voluntarias
· Praderas de plantas residentes útiles
2. Píricas
– Con aporte adicional de disemínulas
– Sin aporte adicional de disemínulas
3. Fertiseres
– Con aporte adicional de disemínulas
– Sin aporte adicional de disemínulas
Praderas en equilibrio
• Clímax o en equilibrio natural con el ambiente
1. Zonales
– Patagónica
· Xérica
· Hídrica
– Puna
· Xérica
· Hídrica
– Veranada de montaña
· Xérica
· Hídrica
2. Azonales
– Salinas
· Salares
· Curso inferior Valles Transversales
– Hídricas no salinas
· Vegas
· Ripiarias
· Ñadis
• Disclímax o sere interrumpida o modificada, pero en equilibrio
1. Zoocenosis
– Especie, raza o tipo de ganado o vida silvestre
– Intensidad y época de utilización
2. Fitocenosis
– Herbicidas selectivos
– Control mecánico selectivo sobre algunos grupos de organismos
3. Edafotopo
– Fertilización mineral y orgánica
– Modificaciones edáficas mecánicas
– Exudados
4. Climatopo
– Microrrelieve modificado
– Modificaciones micro y mesoclimáticas
5. Compleja
Descripción sucesional de las praderas
La comprensión integral del concepto de condición y tendencia requiere una delimitación y definición de los diversos tipos de praderas. Como se trata de un concepto ecológico en su naturaleza, su definición debe corresponder necesariamente a las fases que tratan directamente con aspectos relacionados con los organismos y al medio abiótico en que viven, y de las relaciones intra e interespecíficas de los organismos mismos.
La clasificación ecológica de las praderas requiere, como primera división, la distinción del estado de equilibrio o de evolución sucesional de los componentes biocenósicos, energéticos y de nutrientes que representan las unidades de la pradera, constituyendo los dos grupos principales de ellos.
Las praderas sucesionales son aquellas que se encuentran en cualquier etapa sucesional ajena al clímax en un sentido policlimáxico o en cualquier otra de las etapas sucesionales interrumpidas temporalmente. Ellas, sin embargo, permanecen en forma inducida y mantenida temporalmente por factores controlables que representan desequilibrio. El otro grupo de praderas está representado por praderas en equilibrio, interpretado aquí en el sentido más amplio de la palabra, que significa cualquier estado de estabilidad, sea esta natural o inducida por modificación temporal de algún o algunos factores ambientales, que en condiciones naturales ejercía una distinta magnitud de la intensidad biológica sobre los elementos que constituyen el bioma pratense.
En este sentido “pradera” para Dyksterhuis (1949), se refiere solo a aquellos terrenos cubiertos de pastos nativos en terrenos con pastos naturales o naturalizados de pastoreo. Esta es la acepción tradicional de la palabra y, por lo tanto, la literatura relacionada con condición implícitamente se refiere principalmente a este grupo de praderas. En el presente trabajo se le ha dado una acepción más amplia que incluye tanto a las sucesionales y a las estabilizadas, como asimismo, a las naturales y resembradas. Es preferible pensar, por esta razón, en forma más amplia acerca del clímax, y considerar como tal a cualquier estado de equilibrio sucesional, sea este natural o inducido. En esta forma, la teoría policlimáxica se adapta mejor y más ampliamente al concepto de condición en la forma presentada en este trabajo.
El concepto, en la forma mencionada en este trabajo, es aún más amplio e incluye también a organismos exóticos al sitio, o bien, mejorados genéticamente por medio de cruzas o selección de la población natural, de acuerdo a las necesidades productivas de la pradera. Así se incluye especies de pastos cultivados o pasturas, o bien residentes naturalizados que han sido mejorados en forma antropogénica.
Praderas sucesionales son todos aquellos biomas pratenses, que en un medio físico cualquiera, el componente biocenósico vegetal está compuesto por organismos de corta o larga vida y de corta o larga duración, pero nunca se encuentran estabilizados en clímax, siendo siempre sucesionales. La capacidad de producir evolución sucesional en las fitocenosis que se desarrollan en un sitio es una tendencia natural que conduce, finalmente, a un equilibrio dinámico, lo cual es común a muchas ciencias (Tansley, 1935).
Las praderas sucesionales se dividen en dos grupos principales: los priseres o sucesiones primarias y las subseres o sucesiones secundarias.
Dentro de las praderas priserales, se encuentra uno de los grupos principales representados en el país por las praderas perennes en clímax bosque. Este grupo de praderas representa un área relativamente pequeña en relación al área total donde, la etapa sucesional constituida por pradera perenne se manifiesta durante el desarrollo evolutivo de la sere, cuyo clímax es bosque perennifolio o caducifolio y cuyos componentes principales son árboles de hoja ancha.
La razón de esta singular reducción de la importancia de este grupo característico de praderas se debe a la corta duración en relación al tiempo que demora la sere en llegar al clímax y la duración del clímax mismo. La pradera priseral de pastos perennes pasa casi desapercibida dentro del conjunto evolutivo de la sere y constituye una transición escasamente delimitada. Además, cuando ella se manifiesta, los demás factores del medio ambiente, donde la sere se desarrolla, evolucionan rápidamente y ofrecen condiciones favorables por hierbas altas y gigantes y, frecuentemente, también de arbustos y otras plantas leñosas que pueden normalmente vegetar en esas condiciones. Así, la etapa sucesional representada por hierbas perennes de poco desarrollo, que como productor primario ofrece tejido vegetal útil es reemplazada por otra etapa sucesional más avanzada. Esta evolución progresiva en el desarrollo seral del bioma significa, por lo tanto, una aproximación al clímax. Sin embargo, en relación al bioma pratense significa una evolución retrogresiva o regresión, por cuanto indica un alejamiento de la pradera. La etapa seral óptima es aquella en la cual se obtiene el máximo de tejido vegetal útil, en un sentido pratense.
En praderas sucesionales, en general, sean estas priserales o subserales, retrogresión significa alejamiento de la etapa sucesional pratense más avanzada. En términos pratenses retrogresión pratense puede representar progresión o retroceso. Por esta razón, en el presente trabajo, en lugar de progresión y retrogresión, en el sentido ecológico, se utiliza a menudo el término de alejamiento progresivo y alejamiento retrogresivo, respectivamente. El primero de ellos significa progresión sucesional en un sentido ecológico, pero en etapas evolutivas posteriores a la pradera óptima. El segundo significa retrogresión sucesional en un sentido ecológico, y en sentido pratense significa también retrogresión pratense a partir de la pradera de máximo desarrollo.
Las hidroseres priserales pratenses son, sin embargo, de gran importancia en los climaxes de los bosques templados de lluvia de Chile. A orillas de lagos, ríos y vertientes se encuentran frecuentemente praderas priserales pratenses de alta producción. Sin embargo, en muchas circunstancias es factible argüir que se trata de praderas clímax en equilibrio con el ambiente físico donde se desarrollan. Considerando el equilibrio comunitario en un sentido policlimáxico es posible pensar que se trata de un medio edáfico diferente y, por consiguiente, de dos climaxes edafotópicos diferentes. En muchos casos no ocurre así y se trata simplemente de praderas sucesionales en diferentes etapas evolutivas.
Las praderas de plantas perennes, constitutivas de xeroseres primarios en los climaxes de bosques templados de lluvia son solo de escasa importancia, debido a la pequeña superficie que ellas ocupan.
La pradera sucesional anual en clímax matorral es otro ejemplo de priseres pratenses. Representa un área mayor que la anterior y su existencia se debe a una duración más larga de las etapas sucesionales, en las cuales, las plantas anuales o terófitas representan el mayor componente entre los productores de tejido vegetal útil. Los organismos vegetales anuales endémicos del sitio respectivo se encuentran, a menudo, invadidos e intermezclados con organismos exóticos y naturalizados, los cuales, en conjunto, constituyen las especies residentes. Resulta a menudo difícil encontrar praderas anuales sucesionales, pues la mayor parte de ellas representan algún tipo de clímax. El mayor desarrollo de este grupo de praderas ocurre en la zona del clima mediterráneo del centro de Chile, especialmente en la zona del Norte Chico y provincias administrativas centrales.
Dos grupos de priseres pratenses merecen ser mencionadas en este trabajo. El primero de ellos se refiere a las priseres pratenses psamoserales, que manifiestan su mayor desarrollo en las dunas costeras. Otra prisere de regular importancia es aquella constituida por plantas anuales o terófitas en clímax de pastos perennes. Estas son de escasa importancia por su superficie. Sin embargo, si se considera a la pradera anual mediterránea del centro de Chile, que está constituida por plantas anuales endémicas y naturalizadas, como una etapa sucesional de una sere que concluye en clímax de pastos perennes, este último grupo de praderas sería importante. Sin embargo, debe mencionarse que, en el presente trabajo, la pradera mediterránea anual ha sido considerada como un disclímax o subclímax de clímax matorral o bosque latifoliado de altura reducida.
Las praderas sucesionales son biocenosis intermedias que se desarrollan luego que las poblaciones que constituyen las etapas serales anteriores se autoeliminan y con anterioridad al establecimiento de las poblaciones que invaden y, finalmente, reemplazan a aquellas características de las praderas (Figura 1-4). En ella puede observarse que las poblaciones características de la etapa pratense colonizan y se establecen, presentando tasas de natalidad e inmigración inicialmente altas (), pero ellas decrecen a medida que la interferencia intraespecífica aumenta. Luego y con posterioridad, la tasa de natalidad se incrementa debido a la mayor densidad y, por consiguiente, competencia. Ello es la causa de la invasión de especies características de etapas serales más avanzadas, las que finalmente terminan por dominar el bioma pratense.
Figura 1-4. Esquema hipotético de la variación de la tasa de cambio en praderas sucesionales sin aporte adicional de disemínulas
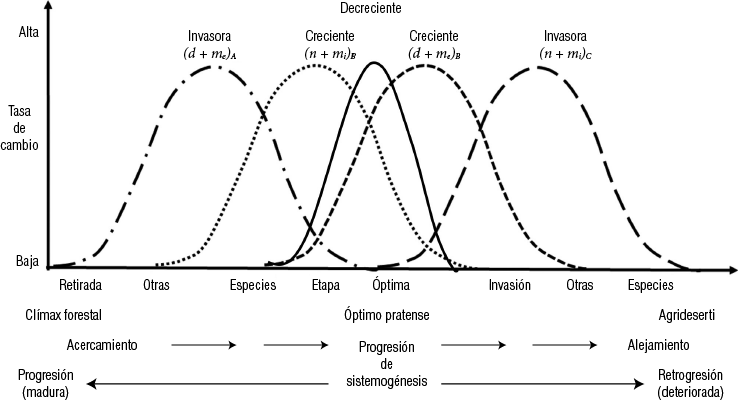
n = natalidad; mi = inmigración; d = mortalidad; me = emigración.
A y C representan especies no características de la etapa óptima, las cuales se presentan, respectivamente, en etapas sucesionales anteriores y posteriores a ella. B representa una especie característica de la etapa óptima (original).
Entre las priseres inducidas cobra especial importancia el control de dunas con Ammophila arenaria, la que posteriormente es reemplazada por bosques naturales o cultivos forestales.
El otro grupo de praderas sucesionales está constituido por subseres pratenses o praderas no climáxicas de las sucesiones secundarias. Uno de los grupos más característicos de praderas pertenecientes a esta categoría está representado por praderas derivadas de la alteración de la cubierta vegetal con implementos que modifican el medio edafotópico, tales como: arados, rastras, rotovator, cultivadoras y otros. Este grupo ha sido denominado cultiseres prístinas o praderas sucesionales prístinas postalteración mecánica.
El grupo recién descrito se divide en dos categorías principales. El primero de ellos, está representado por praderas que necesitan adición de semillas o disemínulas para desarrollarse. En este grupo se encuentran las praderas temporales o anuales, tales como aquellas de avena (Avena sativa); trigo (Triticum aestivum); o cebada (Hordeum vulgare) establecidos para ensilaje, trébol alejandrino (Trifolium alexandrinum), maíz de ensilaje (Zea mayz), sorgo (Sorghum vulgare) y avena con arvejilla (Vicia atropurpurea).
Las praderas bianuales y las de rotación corta que duran más de un año y menos de cuatro, por el hecho de estar constituidas principalmente por especies prístinas, pioneras de las sucesiones secundarias requieren ser mantenidas en las etapas sucesionales iniciales. Así se tiene que tales praderas deben ser destruidas periódicamente con implementos mecánicos y reemplazadas por otros de la misma naturaleza, pero más joven. El objetivo principal de esta práctica es mantener el carácter pionero de los componentes botánicos de la pradera.
La Figura 1-5 presenta un esquema hipotético de la variación de la densidad de las plantas útiles en cultiseres prístinas con aporte adicional de disemínulas y sin reproducción vegetativa posterior a la población aportada. En ella se observa que, al establecer la pradera, la tasa de natalidad y de inmigración es muy alta durante un período relativamente corto, el cual coincide con la germinación de gran número de semillas y la emergencia de las plántulas. Luego, esta línea desciende bruscamente, lo cual coincide con el agotamiento de las semillas potencialmente en condiciones de germinar, pues la reserva de ellas en el suelo es muy baja y el almacenamiento de semillas duras, mucho menor.
Simultáneamente, a la curva anterior se presenta la curva de reducción de la población, debido a mortalidad y emigración. Dada la naturaleza de las especies pratenses se presume que la tasa de emigración es muy baja, cercana a cero y la de mortalidad de plántulas en el período de establecimiento es muy alta. Luego de establecida, la tasa de mortalidad disminuye a un mínimo mientras dura el período de longevidad fisiológica de los organismos que componen la pradera. Cuando la longevidad de los organismos llega a un límite y aparecen organismos fisiológicamente seniles, la tasa de mortalidad de la población aumenta normalmente, al cabo de lo cual la pradera termina por desaparecer.
Figura 1-5. Esquema de la variación de la densidad en pradera culti-sucesionales prístinas con aporte adicional de disemínula y sin reproducción vegetativa

n = natalidad; mi = inmigración; d = mortalidad; me = emigración (original).
Un ejemplo que se ajusta dentro de los límites descritos para este grupo de praderas es el trébol rosado (Trifolium pratense). Se trata de una planta bianual hemicriptófita, que presenta muy mala resiembra natural. Al cabo de uno y medio a dos años la mortalidad de individuos adultos aumenta notablemente y, por lo tanto, la densidad de la especie útil principal disminuye notablemente. La competencia interespecífica originada por Trifolium disminuye, debido al debilitamiento fisiológico de los individuos que la componen como así mismo a la reducción de la densidad. En esta forma, plantas invasoras ocupan los espacios desocupados por los organismos pratenses útiles. El reemplazo ocurre en forma brusca al término del período de longevidad fisiológica de los individuos pratenses que componen la comunidad natural.
La forma de mantener el carácter de prístino de una comunidad compuesta principalmente por individuos pioneros, que no se reproducen en forma natural, es mantener los factores físicos y bióticos del ecosistema en una magnitud biológica similar a los de las comunidades pioneras. Esto se logra con alteración mecánica del medio, ocasionada al término del lapso fisiológico de vida de los individuos que la forman. Además, por tratarse de especies que manifiestan una tasa de natalidad y de migración muy baja, la pradera pionera requiere para su regeneración del aporte adicional de semillas o disemínulas.
La variación de la densidad poblacional de plantas pratenses útiles en praderas cultiserales prístinas que presentan reproducción vegetativa con posterioridad al establecimiento de las plántulas aparece esquemáticamente representada en la Figura 1 6. En ella se resumen las causas de los aumentos debido a natalidad y las etapas características de la mortalidad ecológica y fisiológica.
Figura 1-6. Esquema hipotético, de la variación de la densidad poblacional de las plantas pratenses útiles en las praderas culti-sucesionales, prístinas, con aporte adicional de disemínulas y con reproducción vegetativa post-establecimiento (original)
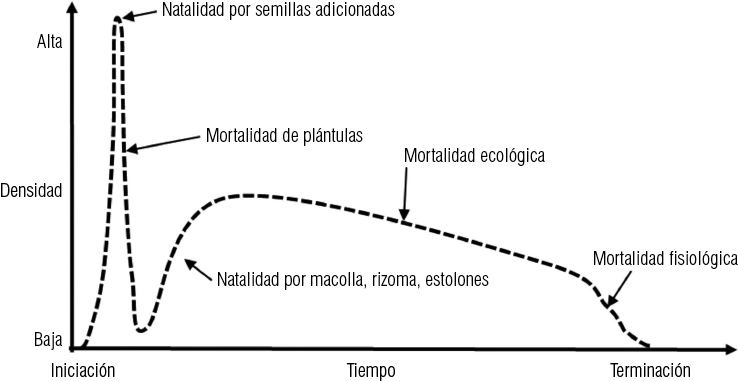
Otras especies ampliamente cultivadas, tales como: alfalfa (Medicago sativa) y ballica inglesa (Lolium perenne), también hemicriptófita, corresponden a este grupo. La mayor diferencia es, sin embargo, que el lapso de vida de los individuos en el medio ecológico natural es generalmente mayor de tres a cuatro años. Al término de este período, la densidad de los individuos en el medio ecológico natural es generalmente mayor, de tres a cuatro años. Al término de este período, la densidad de los individuos pratenses comienza a disminuir debido al incremento de la mortalidad y una natalidad muy baja, nula o casi nula.
Puede ocurrir, sin embargo, que por razones ecofisiológicas, el largo de vida fisiológica de los individuos que constituyen el principal elemento botánico de la pradera sea mayor. Tal es el caso en los valles transversales del desierto de Atacama donde se encuentran algunos alfalfares (Medicago sativa), de diez o más años de edad, posiblemente al no ser afectadas por enfermedades a nivel de la raíz, como ocurre en la zona central de riego. En este caso se trata también de cultisucesiones prístinas. La única diferencia es de magnitud, pues el largo de vida de las plantas es mayor, por lo tanto, la tasa de mortalidad es menor y, además, la tasa de natalidad es, también, algo mayor. La competencia ejercida por organismos vegetales invasores, es menor debido a una flora potencial de especies residentes más pobre. Además, la presencia y acción de fitoparásitos causantes de muerte prematura de plantas, tales como nemátodos del tallo y de la raíz (Ditylenchus dipsaci, Kuhn) es mejor, pues el aislamiento y las condiciones ambientales son menos favorables para el desarrollo de esta enfermedad. En otros casos se produce prolongación fisiológica de la longevidad de los individuos que componen la pradera debido a sus relaciones internas del corte. Tal es el caso de trébol blanco (Trifolium repens), que bajo la influencia del pacimiento presenta tasas de mortalidad más bajas.
El mayor desarrollo de las praderas cultivadas ha sido principalmente dentro de las praderas sucesionales no clímax. Es natural que así haya ocurrido, pues se trata generalmente de plantas de corta vida, altos rendimientos y capacidad competitiva interespecífica muy alta. Por lo tanto, el mejoramiento genético ha sido más rápido y eficiente.
Cualquier tipo de progresión sucesional que ocurra en este tipo de pradera, a pesar de representar un mejoramiento de la condición ecológica para la pradera significa un deterioro, pues ocasiona un alejamiento progresivo y una disminución de la tasa de natalidad de las especies deseables, unido a un incremento de su mortalidad. La única forma de mantener este tipo de praderas es mediante la aradura y resiembra periódica de los principales componentes botánicos del bioma.
Otro grupo importante de praderas está constituido por las mismas praderas culti-sucesionales prístinas postalteración mecánica del medio edáfico, pero en este caso, sin aporte adicional de semilla o disemínulas. Estas son las praderas que crecen después de barbechos que por una u otra razón han sido abandonados sin sembrarse, o bien, después de cultivos tales como: cereales, maíz, legumbres, remolacha y hortalizas. Posterior a la cosecha del cultivo, la tierra es abandonada y no se recurre a la adición de semillas o disemínulas de especies forrajeras o pasturas, son las denominadas praderas rastrojeras (a partir de rastrojos de cultivos). Estas son praderas de malezas o naturalizadas denominadas posíos de primer año o “n” años y generalmente son de gran variabilidad en su composición botánica de acuerdo al sitio y uso previo del área. Generalmente la proporción y el volumen total de forraje útil producido son escasos, aunque en algunos casos es significativo, por ejemplo, al contar con especies como Medicago arabica, entre otras, además de Lolium multiflorum, ambas anuales, voluntarias.
En todos aquellos sitios donde el clímax está constituido por organismos fisionómicamente diferentes a los productores de forraje útil, el alejamiento progresivo de la etapa sucesional pionera, aun en aquellas praderas donde se ha aplicado una adición de semillas o disemínulas ocasiona un deterioro de la pradera.
Pirosucesiones o praderas sucesionales píricas son importantes en zonas de bosques, mata y matorral donde el fuego haya destruido la cubierta natural. La producción de tejido vegetal útil es, a menudo, superior cuando las plantas leñosas no útiles han sido eliminadas. La pradera, que bajo estas condiciones se desarrolla es generalmente más productiva, aun bajo circunstancias donde no se ha hecho un aporte adicional de semillas.
El fuego como elemento inductor de sucesiones suele usarse también en forma exitosa en praderas clímax de pastos perennes. El fuego o roce se repite periódicamente en lapsos cortos de tiempo con el objeto de incrementar la producción de tejido vegetal útil, eliminando el tejido decrépito o lignificado. Así, en esta forma, se modifica la proporción de organismos útiles, aumentando la densidad de los más deseables y disminuyendo la densidad de los pocos deseables. Fuera de ello, la proporción de tejido utilizable proporcionada por los organismos útiles también aumenta, y en esta forma, la biomasa producida de forraje útil en la pradera, aumenta. Quemas periódicas en zonas de bosques y matorral son también frecuentes. Es una de las formas más económicas y simples de aumentar la producción de la pradera, especialmente en aquellos sitios donde la capacidad de uso de la tierra corresponde a pradera. Sin embargo, hoy en día es una práctica no recomendable por los muchos efectos negativos que genera tanto en el suelo como en el entorno donde se efectúa. En praderas pirosucesionales el máximo desarrollo de la capacidad productiva se logra en intervalos muy cortos después de la iniciación de la sucesión secundaria, en tal forma que en la práctica cualquier cambio en la etapa sucesional significa, en la mayoría de los casos, un alejamiento sucesional progresivo sea este en sitios con clímax bosque o matorral y, a menudo, incluso en clímax pradera de plantas perennes.
La inducción de sucesiones secundarias en áreas previamente alteradas por agentes antropogénicos significa generalmente avance pratense. Caminos abandonados, cortes de terrenos y otros movimientos de tierra se manifiestan originalmente descubiertos de vegetación. El cese de la acción humana significa un lento acercamiento sucesional progresivo hacia la pradera y, por consiguiente, un aumento en la productividad de forraje útil. Ocasionalmente, luego que la etapa sucesional pratense óptima ha sido lograda, ocurre un alejamiento progresivo hacia el clímax. Esto significa en muchos casos, el desarrollo de una cubierta vegetal más favorable desde el punto de vista de conservación del suelo, en especial en lo que se refiere a cortes del terreno u otras áreas susceptibles de erosión.
La remoción de vegetación arbórea o arbustiva por medios mecánicos sin la adición de otros recursos bióticos significa también inducción de sucesiones vegetales. Algunas de las etapas sucesionales intermedias representan, frecuentemente, formas fisionómicas características de praderas cuyo tejido vegetal producido es, a menudo, utilizado por consumidores primarios útiles.
Las praderas postdisclímax pratense son aquellas en las cuales se manifiestan sucesiones, pero estas son inducidas por el cese de factores limitantes temporales que han dejado de actuar, los cuales previamente han mantenido un disclímax por un período prolongado de tiempo.
Finalmente, puede mencionarse entre las subseres a las praderas postdisclímax fertilizante o fertiseres. Son aquellas etapas serales retrogresivas que se manifiestan en praderas post clímax que han sido mantenidas en ese estado debido a la adición periódica de elementos fertilizantes aportados desde fuera del ecosistema. Cuando la adición periódica de fertilizantes se interrumpe, se produce una retrogresión hacia el clímax natural, o bien, hacia otro disclímax. Algunas prácticas frecuentes de manejo de praderas están orientadas hacia el desarrollo de praderas sucesionales intermedias de transición desde el disclímax al clímax u otra forma de disclímax.
Las praderas sucesionales, son en sí mismas de una naturaleza tal que por muy bien que se les maneje deben finalmente autodestruirse. La razón de ello es que por no estar en equilibrio están constantemente produciendo modificaciones en su balance hídrico, energético y ciclos biogeoquímicos. Además, aunque sean bien manejadas las tasas de natalidad de las mejores especies son muy bajas y, por lo tanto, no se regeneran a sí mismas.
El avance sucesional significa en este caso empeoramiento de la condición.
Las praderas en equilibrio, al contrario de las anteriores cuando se encuentran sometidas a un régimen de buen manejo, no deben ser resembradas ni regeneradas, puesto que por su naturaleza no producen cambios que conduzcan finalmente a una nueva etapa sucesional.
Las praderas en equilibrio se dividen en dos grupos principales: el primero de ellos se refiere a las praderas verdaderas o clímax. Este grupo es de mayor importancia por cuanto representa el equilibrio natural del ecosistema. El equilibrio pratense de este grupo de praderas significa que los factores físicos y biológicos, animales y vegetales son de tal magnitud biológica que mantienen indefinidamente un bioma que fisionómicamente corresponde a las características pratenses. El segundo grupo representa a las praderas disclímax.
La productividad máxima de la pradera, tanto en relación a su potencial biológico como a las implicaciones económicas que de ella derivan es mayor a medida que se obtiene un acercamiento climáxico. Sin embargo, en forma práctica, a menudo, no es posible mantener el clímax natural por cuanto en manejo ganadero es necesario hacer algunos cambios que directa o indirectamente modifican el ecosistema natural.
Las especies animales que constituyen la mayor causa de consumo del bioma son corrientemente reemplazadas por otras especies domesticadas que transforman el tejido vegetal en compuestos orgánicos de mayor utilidad para el consumidor, en este caso el hombre. Así los diversos organismos herbívoros que en forma natural utilizan la biomasa producida por el bioma son reemplazados por ganado. Los mamíferos herbívoros superiores salvajes, como así mismo otros grupos de herbívoros tales como: aves, insectos, ácaros, nemátodos, etc., son a veces más eficaces transformadores de energía. Sin embargo, a menudo es conveniente reemplazarlos por animales domésticos por cuanto la cosecha posterior de los consumidores primarios se simplifica y, al mismo tiempo, los rendimientos de productos consumibles por el hombre también aumentan. El tamaño de los campos disminuye, el pisoteo y aguadas, la carga animal en proporción a la biomasa utilizable se modifica y otros cambios menores, son los causantes de los cambios resultantes en la pradera clímax.
El concepto de condición ha sido tradicionalmente aplicado a este grupo de praderas. La definición tradicional del concepto así lo indica, ya que condición ha sido definido como el porcentaje de plantas clímax (Dyksterhuis, 1949). Esta definición supone, entonces, que lo mejor debe ser la pradera en equilibrio o clímax, lo cual como se ha señalado en los párrafos anteriores, no es necesariamente una realidad, puesto que alguna etapa sucesional diferente al clímax puede producir mayor biomasa de tejido vegetal útil, ya se trate de biomas en estado sucesional o de equilibrio.
En forma tradicional se ha considerado que el clímax es la etapa sucesional más productiva. Sin embargo, no puede ser así, por cuanto los ecólogos teóricos han demostrado que el clímax Pb – R = Pn = 0. Así la producción neta de biomasa útil por unidad de superficie y tiempo es cero. Lo que se persigue es, por lo tanto, mantener la biocenosis en una etapa anterior al clímax, donde es mayor de cero. La clasificación de la condición se hace, sin embargo, en relación a un patrón de comparación standard y bien conocido para cada sitio, el clímax.
Las praderas zonales son de mayor significación en relación a la superficie que ellas ocupan. Los principales grupos representados en el país son las praderas de hierbas perennes, tales como: la patagónica (Festucetum gracillimae), punense (Festuca ortophilla), veranadas altomontanas (Festuca scabriuscula Phil) y la mediterránea perenne, hemicriptófita (Nasella chilensis, u Hordeun chilense). Esta última, hipotéticamente tuvo importancia en tiempos pretéritos, aunque no ha sido posible demostrar que realmente hubo tal formación en el pasado. Si así ocurriera, el clímax real sería la pradera de especies perennes y, por lo tanto, el matorral y la pradera anual mediterránea serían solo disclimaxes.
Las praderas azonales mantienen su equilibrio limitadas generalmente por algún factor edáfico, principalmente napa freática y sales solubles. Ocupan pequeños sectores a orillas de ríos, lagos y costa, o bien regiones que por su constitución geológica presentan características hídricas locales diferentes de las de otras regiones vecinas.
Las praderas climaxes azonales más prominentes en el país son las que se desarrollan en el curso inferior de los ríos en la zona del desierto de Atacama y Valles Transversales, donde existe una excesiva acumulación de sales.
Entre las hídricas no salinas puede mencionarse a las ribereñas o riparianas que existen a lo largo de los bordes de los ríos y las vegas que se desarrollan en las zonas de emergencia natural de agua (Lotus corniculatus var tenuis). Finalmente, los ñadis que obedecen a circunstancias climáticas y geológicas de la zona sur y que, en algunos lugares, desarrollan praderas (Lotus tenuis), aunque en la mayoría de los casos el bosque es el producto climácico. Las praderas hídricas no salinas pueden ser consideradas como clímax solo si se utiliza el significado policlimáxico de la palabra. Utilizando el significado monoximáxico en la forma tradicional, estas praderas solo representarían una etapa sucesional intermedia de un clímax bosque.
Las praderas disclímax se encuentran en el país bajo las más variadas condiciones ambientales. Representan una proporción muy importante de la superficie total de ellas. El manejo de la pradera en forma tal de mantener indefinidamente un equilibrio sucesional permanente puede representar la obtención de una productividad de biomasa muy alta.
El término “ecosistema antropogénico” fue propuesto por Tansley (1935), para la vegetación que se origina directa o indirectamente bajo la acción de tratamientos culturales del hombre. Bosques transformados en praderas bajo la acción de animales de pastoreo fue sugerido como un ejemplo por el mismo autor.
Las praderas disclímax se caracterizan por presentar un balance de estabilidad de los diversos elementos integradores del ecosistema, ya sea, que se trate de procesos, funciones, niveles u organismos. Se diferencian de las praderas clímax en que la magnitud biológica de los diversos elementos ambientales es diferente a la magnitud natural de ellos, en la forma que se presentan en ecosistemas naturales. De acuerdo al elemento modificado, el cual permite, finalmente, mantener el equilibrio a un distinto nivel que el del clímax se han dividido las praderas en cuatro categorías: zoocenosis, fitocenosis, edafotopos y climatopos.
Los animales pratenses que son parte integral de la zoocenosis son considerados como la causa modificadora del medio pratense. La vegetación natural en equilibrio en suelos zonales es más estable que la vegetación natural desarrollada en equilibrio con un suelo inmaduro; pero el suelo es también considerado como un elemento del ecosistema, lo cual coincide con el punto de vista policlimáxico (Dyksterhuis 1958).
Algunos de los variados beneficios que se pueden obtener en la vegetación debido a la influencia de los animales herbívoros han sido resumidos por Ellison (1960). El ramoneo y pacimiento estimula la producción de flores y frutos. La utilización de la vegetación herbácea puede resultar en una disminución de las reservas radiculares, de follaje, raíces y producción de flores.
El pacimiento de la vegetación puede contribuir a producir modificaciones anátomo-morfológicas de la vegetación que le permita soportar en mejor forma la sequía, al reducir la superficie foliar y establecer una relación más favorable entre la copa y el sistema radicular. Al mismo tiempo, el pacimiento reduce la cantidad de mantillo, lo cual finalmente se traduce en un mejor y más precoz crecimiento de la vegetación pratense.
Los animales son también activos transportadores de disemínulas de especies pratenses, tanto interna como externamente. Al mismo tiempo, mediante el pisoteo contribuyen a situar las disemínulas en condiciones más favorables para la germinación y desarrollo inicial. Los senderos del ganado, que tienden a seguir el contorno de las laderas, detienen el escurrimiento superficial del agua contribuyendo así a una mayor infiltración. Sin embargo, la existencia de huellas bien definidas, es indicativo de pastoreo excesivo, lo cual es, a la vez, responsable de una menor infiltración. Finalmente, una de las influencias mayores de los animales de pastoreo es la producción de deyecciones sólidas y líquidas, mediante las cuales se fertiliza la pradera. Este proceso no significa aporte de nuevos elementos al suelo, puesto que es lo que obtienen de la mayor vegetación que consumen.
Toda la evidencia que se tiene en relación de la dependencia de las especies pratenses con los animales que las utilizan es considerada de naturaleza negativa para el vegetal y positiva para el animal, es decir, un consortismo del tipo parasitario. Sin embargo, toda la evidencia que se tiene indica también que después de milenios de utilización de praderas, la vegetación sigue dominada por las especies más palatables, las que a la larga son beneficiosas. En general, puede concluirse que los beneficios del pastoreo son sobre el ecosistema y la pradera y no sobre las especies palatables o preferidas por el ganado, las cuales son primordialmente consumidas (Ellison, 1960).
La especie, raza o clase de ganado o vida silvestre y la intensidad y época de utilización de la pradera pueden ser modificadas por quien maneje y utilice la pradera. Ello induce sobre el bioma pratense modificaciones de variada naturaleza, los cuales finalmente ocasionan cambios en la composición botánica y por ello en la productividad.
Las fitocenosis pueden, también, ser modificadas mediante la acción directa y mediante el control mecánico selectivo sobre algunos grupos de organismos. La presencia de algunos grupos de plantas, tales como algunas terófitas mediterráneas puede ser la causa de la detención permanente de las sucesiones en un nivel inferior al clímax.
La consideración principal que debe tenerse al mantener un disclímax es determinar si este realmente representa una productividad alta o baja, en relación al potencial del sitio. Diversos disclímaxes pueden producirse en cada sitio y ellos pueden ser más productivos que las praderas sucesionales o de clímax natural. La manipulación de los factores ambientales puede conducir, finalmente, a un incremento de la productividad de biomasa por unidad de área y tiempo.
El avance alcanzado en los últimos años por las ciencias agropecuarias y forestales en el campo tales como: suelos, bioquímica, genética, ecología, fisiología vegetal y animal, nutrición y otros campos que tienen directa influencia en el funcionamiento del ecosistema ha permitido conocer en forma más objetiva la naturaleza de las limitaciones de la productividad. Este conocimiento más profundo, unido, a su vez, a la capacidad de modificar muchos de los factores edafotópicos y climatópicos del medio, permite, en muchos casos, mantener praderas disclímax de mayor productividad que las sucesionales, como las del clímax.
La naturaleza de las relaciones sinecológicas de las praderas disclímax hacen posible utilizar el concepto de condición en la misma forma que se ha utilizado en aquellas de clímax natural. El concepto puede aplicarse en forma similar a lo que se hace con otras praderas. Solo debe determinarse las funciones matemáticas que relacionan las características con la productividad primaria y secundaria del bioma pratense.
Smika et al (1963), por ejemplo, encontraron respuesta diferente de variadas especies pratenses a la acción de los fertilizantes minerales, cultivos y la interacción de ambos. Los pastos nativos y naturalizados no mostraron respuesta a la aplicación de fertilizantes nitrogenados y fosfatados, pero en cambio, Agropyron desertorum, aumentó, al mismo tiempo que redujo el porcentaje de malezas en la pradera, pues debido a la acción del fertilizante se aumentó la competencia con las malezas, como resultado del mayor desarrollo y crecimiento alcanzado por esta gramínea. Esto conduce, finalmente, a diferentes cauces sucesionales que se generan a distintas velocidades y conducen a otros disclímaxes.
Las especies pratenses reaccionan diferentemente al pastoreo o a la remoción de tejido, en general. En ciertas épocas del año se observa una reducción de la producción de semillas de algunas especies, más que de otras, lo cual limita la prevalencia de algunas especies en la vegetación (Horton et al, 1957).
Las leguminosas de raíces profundizadoras (Medicago sativa L) almacenan sus reservas de carbohidratos en las raíces, pero las estoloníferas (Trifolium repens) lo hacen en los estolones, en todos aquellos casos la altura de corte no es responsable de la remoción de carbohidratos con el follaje. En gramíneas, en cambio, los carbohidratos no estructurales se almacenan en parte en las raíces y rizomas, pero primordialmente en el sector inferior de la vaina y, por lo tanto, al corte o pacimiento muy bajo es responsable también de una remoción considerable de los carbohidratos no estructurales almacenados en la planta. La velocidad de recuperación y el vigor se reduce al ser sometidos a cortes o pacimientos muy bajos. Para las gramíneas, la altura de corte no es tan crítica como lo es la frecuencia y época de defoliación (Sprague, 1952).
Reed y Peterson (1961), llegaron a la conclusión que, bajo la presión ejercida por el pastoreo estival más intensivo, se reduce la altura de los culmos florales de las gramíneas forrajeras. Además, se reduce el peso de los culmos, lo que se traduce, finalmente, en la disminución de la producción de las especies más palatables. Simultáneamente, ocurre un predominio de las especies de menor altura, las cuales, a su vez, son menos accesibles al ganado vacuno.
En regiones de pastos altos, bajo condiciones de pastoreo intensivo, se ha visto disminución de las especies que componen la estrata herbácea superior y, al mismo tiempo, dan cabida a pastos cortos, especies anuales y leñosas. Dyksterhuis (1946), atribuye a la presencia e invasión de árboles y arbustos formando sabanas y matorrales a la reducción de la competencia por la humedad del suelo, que se produce al sobrepastorear la pradera, debido a la disminución del desarrollo de la copa y sistema radicular de los pastos palatables.
El grado de perfección de un ecosistema pratense clímax se mide por la relativa estabilidad del equilibrio entre clima, vegetación y suelo (Dyksterhuis, 1958), y por la integridad de la utilización de los componentes ecotrópicos y biocenóticos al nivel más alto.