Las raíces profundas
2
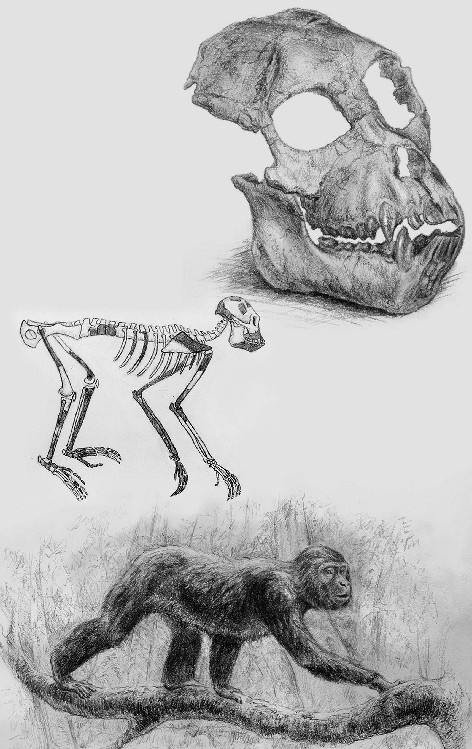
En el lado keniata del lago Victoria, en una isla llamada Rusinga, Louis y Mary Leakey efectuaron en 1948 uno de sus muchos y relevantes hallazgos. Junto con otros restos de mamíferos de edad miocena, encontraron el cráneo y algunos huesos de las extremidades de un antiguo simio al que llamaron Proconsul, un género de primates cuyo apelativo hacía honor a Cónsul, un famoso chimpancé del zoológico de Londres. La repercusión del hallazgo fue grande y desde entonces se han ido recuperando otros muchos fósiles en más de seis yacimientos de la citada isla, hoy datados en nada menos que hace 23 Ma.
La antigüedad y las circunstancias hicieron ver en Proconsul al representante del tronco común de primates del que debieron de surgir los simios y los humanos. El impacto teórico de esta aseveración fue enorme, ya que se trataba de un simio arborícola pero de locomoción cuadrúpeda y con un cuerpo de constitución muy similar al de los micos. No se detectaron en él los caracteres anatómicos clásicamente asociados a la suspensión bajo ramas, propia de los simios actuales, un hecho decisivo que vino a contradecir el modelo «braquiador», implantado desde inicios del siglo XX, con el que se pretendía explicar el precedente humano. Este episodio es uno de los más complejos en el estudio de la evolución humana y merece una buena explicación.
LA SUSPENSIÓN BAJO RAMAS
Como ya hemos señalado, los científicos del siglo XIX, y en especial Darwin, centraron su teoría en un pasado arborícola prehumano y consideraron que el bajar de los árboles fue el paso inicial para el proceso de humanización. Pero ¿fue realmente el UAC un animal arborícola? De ser así, ¿cómo vivía y se desplazaba por el dosel de la selva? ¿Qué locomoción practicaba para poder adoptar después un andar bípedo? En la anatomía del cuerpo humano están las respuestas de una vida en los árboles.
En 1903 aparece en escena el anatomista y antropólogo escocés Arthur Keith, quien será el primero en proponer una hipótesis bien definida sobre el tipo de locomoción arbórea que precedió al bipedismo. Sobre la base de sus observaciones en los bosques de Siam (Tailandia), adonde se dirigió para ejercer como médico de una compañía minera, y sobre todo a partir de las disecciones de macacos, langures y gibones, comprobó que las conductas locomotoras de los simios en los árboles tenían mucho que decir sobre cómo se había llegado a la postura erguida. Según Keith, todos los hominoideos compartimos la arquitectura especial del torso: ancho y plano, e íntimamente relacionado con una postura ortógrada. Este rasgo básico está a su vez vinculado con una clavícula larga y una orientación lateral de la cavidad glenoide (donde se inserta la cabeza del húmero). Para Keith y sus seguidores, los primates hominoideos presentan una adaptación locomotora relevante: la braquiación, es decir, desplazarse balanceándose entre las ramas y valiéndose del uso alterno de las manos (aunque actualmente se entiende por braquiación un modo particular de locomoción casi exclusivo de los hilobátidos [gibones]).
Si nuestros familiares evolutivos más próximos presentan la adaptación braquiadora, cabe suponer que el UAC también la conservaría. La verticalización del tronco derivada de la suspensión del cuerpo bajo las ramas era el modo de vida de los prehumanos y, consecuentemente, el precedente de la locomoción bípeda. En el esquema teórico del anatomista escocés, la fase en la que se establecieron las bases anatómicas de la postura ortógrada y la capacidad de suspensión fue la llamada hilobátida, esto es, un pequeño primate ortógrado* hilobátido, surgido por el cambio de un mono pronógrado* cuadrúpedo arborícola (que se desplaza sobre las ramas) a un simio braquiador (que se cuelga de las ramas). Los ancestros de gibones, grandes simios y humanos adquirieron entonces largos brazos, muñecas con amplia capacidad de giro, hombros muy móviles y torsos anchos y planos. Una segunda fase, llamada troglodítica, resultaría en un incremento en el tamaño del animal, similar a lo observado en chimpancés y gorilas, cuyos efectos serían determinantes para que finalmente surgiera el linaje humano.
Un grupo de simios arborícolas gigantes se adaptó a una locomoción plantígrada en el suelo como consecuencia, en gran medida, de la dificultad de una vida arbórea por su gran talla (los gorilas actuales son un buen ejemplo de esto: pasan buena parte de su vida en el suelo, especialmente los machos adultos). El modelo de braquiación plantea, pues, que los simios y los humanos comparten una forma del cuerpo erguida (ortógrada) como resultado de una herencia común de adaptación a la suspensión en las ramas. El tamaño y la disposición de las piernas, extendidas y colgantes, que ayudan al balanceo, darían una anatomía preadaptada para un posterior bipedismo terrestre.
Ciertamente el modelo de Keith tuvo una gran capacidad de seducción. El escenario del torso erguido para llegar al bipedismo encajaba con el necesario transporte de herramientas y la concomitante encefalización del modelo darwinista. (Recordemos que la teoría ecológica de causación de Darwin distingue cuatro grandes eventos en la evolución humana: 1) el origen de la terrestralidad [bajar de los árboles], 2) la postura erguida y la liberación de las manos, 3) la encefalización y 4) el surgimiento de la cultura [civilización, según algunos].) La hipótesis de una fase braquiadora en la ascendencia humana dominaría la escena académica y popular durante décadas, pero el hallazgo de los fósiles de la isla de Rusinga hizo colapsar este modelo. Como ya señalamos, Proconsul tenía un cuerpo de mono pronógrado (torso inclinado hacia delante) que no respondía a las predicciones de simio de torso erecto. Aunque hoy sabemos que la antigüedad de Proconsul no tendría por qué invalidar los razonamientos de Keith, este hecho ayudó, junto con otras evidencias, al rechazo de su teoría.
LOS RASGOS PRIMITIVOS DEL CUERPO HUMANO
Ante el modelo que veía al antepasado humano con un cuerpo construido según el bauplan* de los simios actuales —especializaciones anatómicas que se manifiestan sensiblemente en el modo de locomoción—, otros situaban nuestras raíces antes de la aparición del esquema corporal de los simios. Desde ángulos diferentes se sostenía que el origen humano, y con él el bipedismo, arrancaba de un grupo de primates en el que todavía no se habían expresado con claridad las características de los simios, y con ellas la braquiación. Esto suponía que humanos y antropoides habían surgido independientemente, y por eso el cuerpo humano preserva rasgos primitivos que se han perdido en la evolución de los grandes simios.
Para el naturalista americano Willian Straus, la mano humana es de una anatomía generalizada muy similar a la de los cercopitecoideos, lo que le llevó a proponer que los humanos se habían originado desde un ancestro similar a los micos de locomoción sobre ramas. Asimismo, los primatólogos Adolph Schultz y Le Gros Clark secundaron la teoría de que los humanos proceden de un acervo común con los simios, pero anterior a que se desarrollasen las peculiaridades de estos. Bajo la influencia de Le Gros Clark nació una corriente centrada en la idea de que orangutanes, gorilas y chimpancés presentan caracteres muy especializados que no se reconocen en nuestra anatomía, y hasta el propio Keith se avino finalmente a un origen prebraquiador para el linaje de los homininos.
Según esta perspectiva, los grandes simios formaban un grupo taxonómico propio, mientras que la singularidad de los humanos definiría otro grupo bien diferenciado. Tal distancia genealógica planteaba unas hondas raíces evolutivas para el linaje humano y, por lo tanto, un largo periodo desde la divergencia con los simios, que se remontaría cuando menos al Mioceno. Los fósiles del antropoide mioceno Dryopithecus encontrados en Europa proporcionaron una evidencia paleontológica a este modelo. Fue en este contexto, y más concretamente en el año 1927, cuando Pilgrim formalizó la separación taxonómica de las familias zoológicas de los póngidos y los homínidos, la cual estuvo vigente durante décadas.
UN FALSO FÓSIL: LA VERGÜENZA DEL GREMIO
En 1912 se hizo público el hallazgo de unos restos en Sussex (Inglaterra), que se presentaron en la Sociedad Geológica de Londres con el nombre de Eoanthropus dawsoni. Se trataba del famoso hombre de Piltdown, identificado a través de fragmentos de un cráneo de aspecto y tamaño propios de un humano, junto con una mandíbula de anatomía muy primitiva. Los fósiles de mamíferos de la misma capa estratigráfica de donde procedían parecían remitir su antigüedad tan atrás como el Plioceno.* En la época fue reconocido como el eslabón perdido entre los simios y el hombre, y adquirió verdadera notoriedad en la literatura científica y la prensa.
En realidad eran los restos de un cráneo humano y un fragmento de mandíbula de orangután actual convenientemente tratada y teñida como para pasar por un fósil, a los que se sumaron después unos supuestos dientes de chimpancé. En 1953, más de cuarenta años después, la revista Time publicó los datos de científicos ingleses que desvelaban la naturaleza fraudulenta del caso. En su momento hubo expertos que dudaron de la validez del hallazgo, pero ¿qué llevó a otros a aceptarlo? Una mezcla de circunstancias, desde la competencia en la búsqueda del eslabón perdido por parte de Inglaterra y países como Francia y Bélgica hasta el escaso acceso de los investigadores internacionales al estudio del material, pero sobre todo el hecho de que Piltdown venía a demostrar, a su manera, las ideas de Darwin.
Los «restos» de Sussex, una vez aceptados por la oficialidad, generaron dos conclusiones. La primera es de índole teórica: se dedujo que, de los rasgos básicos del modelo de Darwin, el incremento del cerebro fue el que inició el proceso de diferenciación entre los seres humanos y el resto de los animales, y esto repercutió después en la aceptación de Australopithecus, que marcará para siempre el rumbo de la paleontología. La segunda conclusión fue que las raíces del linaje humano eran muy antiguas, pues la aparicición de rasgos modernos previa a la diferenciación de los rasgos simios remitía necesariamente a un tiempo mucho más lejano. En este sentido, Piltdown supuso a la vez un retroceso y un avance. Y como avance se encuentra el haber puesto en un primer plano la importancia del registro fósil para entender la evolución humana. Sin duda, a partir del supuesto antepasado Eoanthropus dawsoni, los verdaderos fósiles tuvieron mucho que decir.
LOS ANTEPASADOS DE LOS HOMININOS
Hasta la década de los años sesenta del siglo XX, se pensaba que los primeros representantes de la familia humana tendrían que provenir del Mioceno, hace unos 15 Ma. Los fósiles del simio Ramapithecus, procedentes de los montes Sivalik, en Pakistán, fueron la base de la candidatura propuesta por el paleontólogo americano Edward Simons como representante de la raíz miocena del linaje humano. La forma de su arcada dentaria, el patrón de desgaste y el grosor del esmalte, rasgos compartidos con los humanos modernos, avalaron esta interpretación.
Durante un tiempo se unió a la candidatura el también mioceno Kenyapithecus, encontrado por L. Leakey en Kenia, fósil al que el paleoantropólogo británico proclamó como hacedor de las herramientas prehistóricas que él iba encontrando en diferentes yacimientos keniatas y, por lo tanto, una clara evidencia de la gran antigüedad del linaje humano. Sin embargo, en 1963 este paradigma se resquebraja. Ese año entran en acción las técnicas moleculares, que cambiarán radicalmente el escenario de la evolución humana. La recién fundada antropología molecular reveló una mayor afinidad de las proteínas sanguíneas humanas con las de los simios africanos y proporcionó las primeras estimaciones en los tiempos de divergencia. La fuerza de la biología molecular impuso su criterio: los primates más próximos a los seres humanos son los simios africanos y su divergencia evolutiva es muy reciente (hoy admitimos que es de hace 6 Ma). Desde entonces, una larga lista de resultados genéticos ha venido a confirmar la estrecha relación entre los humanos y los simios africanos, lo que ha ratificado el paradigma de Huxley.
Los datos moleculares cuestionaron de raíz los pretendidos orígenes miocenos de humanos y simios. A esto contribuyó sin duda el descubrimiento en Turquía y Pakistán de nuevos fósiles del simio mioceno Sivapithecus, de unos 8 Ma de antigüedad. Los rasgos del cráneo y en especial de la cara hablaban con claridad de una relación filética con el idiosincrático orangután, a la vez que permitían subsumir los restos de Ramapithecus en esta especie. El candidato a primer representante del linaje humano se reubicaba ahora con toda claridad en la ascendencia de los grandes simios asiáticos. Pero, una vez separados los asiáticos y los africanos, el problema central recayó, como hemos señalado, en las relaciones entre los gorilas, chimpancés y humanos.
LA HIPÓTESIS DE LA TREPA VERTICAL
En la década de los setenta, a medida que los estudios de anatomía comparada se fueron haciendo cada vez más precisos, y fueron abarcando grupos más extensos de animales, se observó que caracteres que se habían asociado a la braquiación aparecían también en animales no braquiadores. El creciente examen de primates en sus medios naturales permitió constatar que sus movimientos eran realmente versátiles, y no resultaba nada fácil encasillarlos en patrones estancos. El chimpancé, por ejemplo, practica en el suelo una locomoción cuadrúpeda —el nudilleo—, pero en los árboles se vale de una variedad de modos locomotores, desde la trepa vertical a la suspensión y balanceo bajo ramas, por no hablar de los cambios locomotores que suceden a lo largo de su periodo de crecimiento. Por otro lado, el uso de nuevas técnicas, como la electromiografía —la medición de la acción de los músculos en determinadas acciones—, completó el panorama de revisión y mostró que, cuando los simios trepan (y también los atelinos, unos monos sudamericanos que practican la braquiación), ponen en acción los mismos músculos del brazo que utilizan en la suspensión. En definitiva, los avances técnicos sugerían que la anatomía de los brazos podía ser mejor explicada como una adaptación a trepar en soportes verticales, y por eso a inicios de los años setenta del siglo XX se gestó la idea de que la acción de trepar, antes que la braquiación, era la clave para entender el paso de una vida arborícola a una terrestre.
Con la aplicación de estos nuevos métodos, uno de los aspectos que más llamó la atención fue la similitud biomecánica entre la trepa y el bipedismo, puesto que en ambos ejercicios el cuerpo usa los mismos conjuntos musculares. La trepa vertical se consideró entonces como preadaptativa para los movimientos de las piernas propios del bipedismo humano. Para subir los troncos de los árboles, los grandes primates usan sus largos y poderosos brazos, piernas cortas y pies prensiles, con grandes dedos gordos. ¿En qué escenario ecológico los chimpancés desarrollaron la adaptación de trepar básicamente con los brazos?
En las selvas tropicales, la luz es uno de los recursos más valorados. Como las plantas se solapan, la competencia por captar fotones es total. La necesidad de alcanzar la luz lleva a muchos árboles a desarrollar troncos muy largos y verticales, desprovistos de ramas laterales. Solo en el estrato superior se ramifican y fructifican. Alcanzar este estrato exige unas habilidades claras para trepar. Al parecer, por adaptación a estos medios, las manos de los simios muestran un gran alargamiento, y su conexión con los huesos de la muñeca está muy fortalecida. El pie, por su parte, acentúa la capacidad de agarre.
En los modelos clásicos de Keith y sus seguidores, la trepa y suspensión no se distinguían y, por lo tanto, se consideraban un único fenómeno evolutivo. Desde los años setenta, la arquitectura corporal se desdobló: por un lado estaban los rasgos asociados a una adaptación más básica y común de los simios (la trepa); por otro, los caracteres más especializados (necesarios para la suspensión bajo ramas).
EL NUDILLEO COMO FASE DE TRANSICIÓN
En la búsqueda posdarwiniana de los precursores inmediatos prehumanos, se produjo una bifurcación teórica con dos dimensiones entrelazadas: por una parte, el modelo de locomoción (braquiación o no); por otra, la antigüedad de la divergencia (remota o reciente). Aunque el modelo braquiador de Keith se desmontó, en cierta medida evolucionó en diferentes direcciones.
En los años cincuenta, el antropólogo estadounidense Sherwood Washburn, a través de la evidencia molecular que recupera la proximidad entre humanos, gorilas y chimpancés, y de estudios morfológicos del esqueleto poscraneal, revitalizó el paradigma mixto Huxley-Keith (a saber, proximidad con simios africanos y braquiación), pero con un formato actualizado. Mientras que el modelo original de Huxley-Keith no reconocía ninguna etapa intermedia entre la braquiación y el bipedismo, la teoría de Washburn incorporó una fase de locomoción terrestre intermedia, similar a la que vemos en los gorilas y chimpancés actuales. Se trata del llamado nudilleo —desplazarse con el apoyo de los nudillos—, y su inclusión como una fase de la evolución representa un nuevo elemento de complejidad. Será con el hallazgo de Ardipithecus cuando se profundice en esta hipótesis.
UNA VISIÓN PALEOGEOGRÁFICA DE LOS SIMIOS
La distribución geográfica de los grandes simios hoy se limita a parcelas aisladas en las selvas lluviosas de África e Indonesia. Los chimpancés y gorilas perviven en los restos del cinturón ecuatorial africano, mientras que los asiáticos orangutanes lo hacen en las islas de Java y Borneo. Esta limitada distribución es reflejo del pasado, y los fósiles nos enseñan que los simios tuvieron durante el Mioceno una distribución geográfica mucho más extensa.
El origen evolutivo de los grandes simios podemos ubicarlo a finales del Oligoceno, hace poco más de 25 Ma, en lo que fue Afroarabia, una masa continental continua antes de que se abriera el mar Rojo. Las primeras fases de su evolución (géneros Proconsul, Afropithecus, etcétera) estuvieron limitadas a este ámbito geográfico, pero a partir de hace unos 16 Ma se produjo una fuerte expansión. Durante el Mioceno medio, diferentes y variadas especies de simios vivieron desde el centro de Europa (Griphopithecus, Dryopithecus, etcétera) hasta el Asia oriental (Sivapithecus), además de la parte central de África (Kenyapithecus). Toda esta expansión está documentada en diferentes yacimientos y con fósiles diversos.
Un magnífico ejemplo de abundancia y diversidad lo encontramos muy cerca de casa, en la cuenca del Vallés-Penedés, donde se han recuperado abundantes fósiles de mamíferos y primates hominoideos (Pierolapithecus, Hispanopithecus, Anoiapithecus). Si observáramos un mapa del planeta Tierra de hace 12 Ma, veríamos una amplia superficie habitada por simios diversos, extendida en un ancho cinturón que abraza por completo el viejo mundo desde el sur de África hasta Centroeuropa, y que abarca selvas y bosques en condiciones ecológicas diversas. A partir de hace 11 Ma se produjo la «crisis mesiniense», lo que generó, entre otras consecuencias, una retracción drástica de las áreas de distribución de los simios. Desde ese momento desaparecen en Europa y Turquía, con la excepción de la pervivencia de algún simio relicto (Oreopithecus) en las antiguas islas del Mediterráneo, Córcega y lo que es hoy la Toscana.
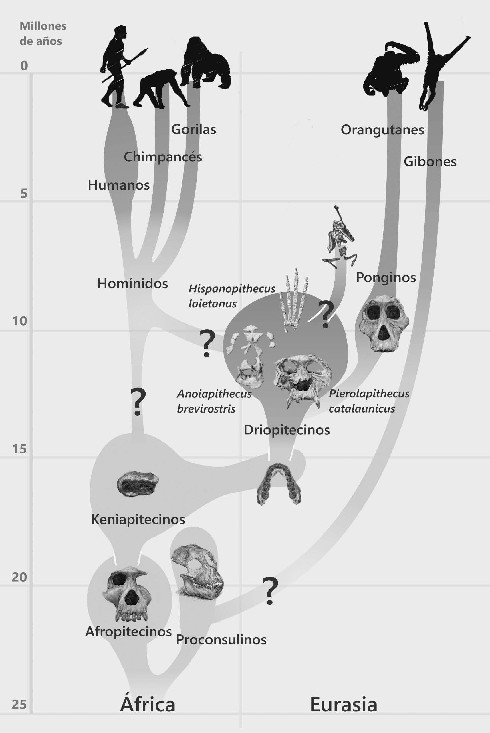
Filogenia de los hominoideos fósiles y actuales.
En el otro extremo de la Tierra hay también supervivencia de especies en el sudeste asiático (Gigantopithecus), lo que posteriormente terminará siendo el área de distribución del orangután. Sabemos que el simio rojo ocupó amplias zonas en el continente asiático hasta hace bien poco. Actualmente los orangutanes malviven en libertad solo en Java y Borneo, además de habitar en algunos parques zoológicos. Con una edad de 8 a 10 Ma, aparecen en Grecia y Turquía restos de un gran simio (Ouranopithecus), cuya anatomía converge con la de los simios africanos. Esta circunstancia ha alimentado la hipótesis no darwiniana de que los homínidos (grandes simios + humanos) evolucionaron en la amplia zona subtropical de Eurasia sobre un esquema corporal dryopitecino. Una posterior dispersión durante el Mioceno superior llevó al clado pongino al sudeste asiático y al clado hominino de vuelta a África (en un evento out-of-Europe).
En el continente africano, aunque el registro fósil de los grandes monos antropomorfos se hace muy fragmentario a partir del Mioceno medio, el descubrimiento reciente de nuevos simios miocenos (Nakalipithecus, Samburupithecus y Chororapithecus) permite enlazar el clado africano con sus orígenes locales remotos. Pero a partir de hace unos 8 Ma de años perdemos casi por completo el rastro al linaje de los simios africanos y curiosamente empieza a hacerse cada vez más abundante el registro paleontológico de una variante de simios bípedos.
En términos generales, el deterioro de los ecosistemas, dominados cada vez más por el avance de la aridez, llevó a los antropoides africanos a una profunda crisis, patente en una considerable merma tanto en su diversidad como en el número de sus miembros. Un linaje de estos simios africanos, cuyos supervivientes actuales son los gorilas y los chimpancés, logró acantonarse en ambientes aún favorables y más estables, a resguardo también de la creciente competencia con los emergentes micos africanos o monos cercopitecoideos (mangabeis, cercopitecos, babuinos, etcétera). De entre este grupo de simios en horas bajas emergió un linaje que experimentó con nuevas adaptaciones, lo que le permitió prosperar en número y diversidad, así como extenderse de nuevo más allá de los confines de África, esta vez en forma bípeda. Este linaje es el de los homininos.
LA AUSENCIA DE FÓSILES
¿Por qué abundan los fósiles de homininos en el este de África en comparación con los escasísimos restos de otros simios? Para explicar la desproporcionada cantidad de restos de estos dos grupos se han esgrimido causas tafonómicas ligadas a los nuevos ambientes ecológicos que ocuparon los homininos, en los que la fosilización era más fácil. A su vez, la formación del Gran Valle del Rift generó altas tasas de sedimentación que favorecieron el enterramiento rápido y, por ende, la fosilización. Sin embargo, paradójicamente nada sabemos del linaje de los chimpancés; apenas un par de dientes de edad pleistocena completan su registro fósil.
Del mismo modo, el hallazgo de fósiles en el África central y occidental es prácticamente inexistente. Apenas hay datos de la inmensa cuenca del Congo o las regiones del golfo de Guinea. Las razones esgrimidas para explicar tal ausencia reposan en las desfavorables condiciones de preservación de restos orgánicos en ambientes selváticos. En este sentido, hay dos vertientes inéditas de la evolución humana (el registro de simios y el registro del oeste de África) que están esperando que vayamos y busquemos esas evidencias paleontológicas, pues las circunstancias de la dicotomía entre Pan y Homo no se podrán resolver hasta que no dispongamos de la documentación que acote los parámetros involucrados en este modelo.
Este es uno de los grandes retos de la paleoantropología actual. El hallazgo de fósiles del Mioceno tardío ha despertado el interés por la reconstrucción del UAC Pan-Homo, uno de los aspectos más interesantes de la teoría de la evolución humana. Cuando encontremos fósiles en las selvas del África occidental todo cambiará.